
|
|
|
|
Бехтерев В. М.
Избранные произведения
(СТАТЬИ И ДОКЛАДЫ)
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
МЕДИЦИНСКОЙ ЛИТЕРАТУРЫ
МЕДГИЗ — 1954 — МОСКВА

Вступительная статья и примечания
профессора В. Н. МЯСИЩЕВА
В.М.БЕХТЕРЕВ
(1857— 1927)
Научная, врачебная и общественная деятельность Владимира Михайловича Бехтерева протекала в последней четверти XIX и первой четверти XX века. В это время в России быстрыми темпами развивается капитализм. Одновременно зарождается и быстро нарастает революционное движение рабочего класса, приведшее под руководством коммунистической партии к победе Великой Октябрьской социалистической революции.
Ученые конца XIX и начала XX века разделились на две группы — прогрессивную, преследуемую царским правительством, и реакционную, поддерживаемую им. В. М. Бехтерев принадлежал к группе прогрессивных ученых.
Будучи прежде всего выдающимся невропатологом и психиатром, В. М. Бехтерев не ограничивал свою деятельность этими областями. Он вел широкие научные исследования в области анатомии и физиологии нервной системы, а также психологии. С самого начала своей научной работы он развернул широкую общественную и организаторскую деятельность, преимущественно в области организации медицины и народного образования.
В связи с 25-летием со дня смерти В. М. Бехтерева проф. А. Г. Иванов-Смоленский писал, что он «является одним из самых выдающихся представителей отечественной психоневрологии, оставившим после себя сотни научных работ, среди которых ряд имеющих фундаментальный характер. Он оставил глубокий и плодотворный след в невропатологии, психиатрии, нейроморфО' логии, психотерапии, в учении о локализации функций, гигиене нервной системы, нейрохирургии, психологии и т. д. Его научные труды получили мировую известность и завоевали в свое время огромный авторитет, преимущественно у клиницистов. Велико также значение В. М. Бехтерева как общественного деятеля и организатора научных учреждений» х.
Владимир Михайлович Бехтерев родился в селе Сорали Ела-бужского уезда Вятской губернии 20 января 1857 г.2. Девяти
1 Журнал высшей нервной деятельности, 1953, т. II, ч. 6.
2 По данным послужного списка. В других источниках указаны различные даты — ст 20/1 до 1/П 1857 г.
3
лет он потерял отца, и семья, состоявшая из 5 человек—матери и четверых сыновей, испытывала большие материальные трудности. Однако умная и культурная мать с большим трудом все же обеспечила детям хорошее образование.
Владимир Михайлович учился в Вятской гимназии. С детства он любил природу и обнаруживал большой интерес к естествознанию, читал с увлечением естественно-научную литературу и, в частности, 'труды Дарвина. В юности он познакомился с произведениями революционных демократов-материалистов— Н. А. Добролюбова, Д. И. Писарева и других, оставившими глубокий след в его теоретических и общественно-политических взглядах.
По окончании гимназии В. М. Бехтерев поступил в Медико-хирургическую, позже Военно-медицинскую, академию в Петербурге. Здесь он с большим увлечением занимался естественными и медицинскими науками, а на четвертом курсе сосредоточил свое внимание на клинике нервных и психических болезней.
В студенческие годы Владимир Михайлович участвовал в выступлениях студентов, демонстрациях рабочих, например, в вошедшей в историю революционного движения в России демонстрации на Казанской площади в декабре 1876 г.
В период весны и лета 1877 г. В. М. Бехтерев прервал свои занятия в Медико-хирургической академии и отправился на театр военных действий на Балканы. В корреспонденциях, которые он присылал оттуда в периодические издания, он отразил свое глубокое преклонение перед героизмом русского солдата и горячую симпатию к славянским народам.
В 1878 г. В. М. Бехтерев окончил Медико-хирургическую академию с отличием и премией и был оставлен при академии для подготовки к профессорской деятельности.
Свою научную работу В. М. Бехтерев начал под руководством И. П. Мержеевского. В 1880 г. он защитил диссертацию на тему «Опыт клинического исследования температуры тела при некоторых формах душевных болезней», в которой психические нарушения и изменения температуры тела рассматривает в единстве, как проявления болезни мозга. Эта диссертация явилась продолжением исследований И. П. Мержеевского, изложенных в работе «Клинические исследования неистовых больных (vesanici)». В этой работе заметно влияние идей С. П. Боткина, заложившего основы учения нервизма в медицине. Вскоре после защиты диссертации В. М. Бехтерев получил звание приват-доцента и был избран профессором Казанского университета.
В Казани Б. М. Бехтерев развернул широкую научную и научно-общественную деятельность. Он превратил окружную психиатрическую больницу в базу психиатрической клиники, используя ее для педагогических и научных целей. Научную работу в области невропатологии он развернул в Казанском военном госпитале. В Казанском университете В. М. Бехтерев
4
устроил первую в России психофизиологическую лабораторию. Из кафедры, руководимой В. М; Бехтеревым, вышло много научных трудов, она привлекала к себе большое число сотрудников. По инициативе В. М. Бехтерева в Казани было организовано общество невропатологов и психиатров, которое начало издавать журнал «Неврологический вестник».
Казань в этот период являлась одним из важнейших культурных центров России. В. М. Бехтерев работал там в тесном содружестве с замечательными учеными, медиками и естествоиспытателями — Н. А. Виноградовым, Н. О. Ковалевским, Е. В. Адамиком, К- Н. Арнштейном, А. М. Зайцевым и др. Особенно тесен был научный контакт его с крупным казанским физиологом Н. А. Миславским, совместно с которым В. М. Бехтерев провел ряд важных исследований.
В. М. Бехтерев содействовал привлечению в Казань на кафедру невропатологии впоследствии одного из виднейших невропатологов Л. О. Даркшевича.
Казань в то время была немаловажным участком пробуждающегося революционного движения; в 80—90-е годы революционная пропаганда казанских народовольцев охватила много рабочих. Создавались нелегальные типографии. В студенческие народнические кружки проникали идеи марксизма. Н. Федосеев в 1888 г. сформировал первый марксистский кружок, в котором участвовал В. И. Ленин.
В. М. Бехтерев привлекал к себе на работу прогрессивно настроенную молодежь. Так, его ученик П. А. Останков, подвергавшийся аресту за участие в студенческих беспорядках, с трудом, лишь при активной поддержке Владимира Михайловича, получил утвержденную в штате кафедры должность; В. М. Бехтерев никогда не сообщал о студентах, отсутствующих на его лекциях вследствие участия в сходках. Уже в этот период В. М. Бехтерев считался неблагонадежным и его не награждали при царском режиме, хотя его научная и педагогическая деятельность была исключительно продуктивной.
В Казани В. М. Бехтерев работал 8 лет. За это время он выпустил двухтомный труд «Нервные болезни в отдельных наблюдениях», а также подготовил первое издание своего классического труда «Проводящие пути спинного и головного мозга».
В 1893 г. В. М. Бехтерев начал работать в Петербурге в Военно-медицинской академии. Несколько позже он был избран также на вновь открывшуюся кафедру нервных и душевных болезней Женского медицинского института. В этот период размах его педагогической, научной, научно-организаторской и научно-общественной деятельности значительно расширился. В Военно-медицинской академии он реорганизовал психиатрическую клинику и построил новую клинику нервных болезней, открыв при ней одно из первых в мире нейрохирургическое отделение, большое физиотерапевтическое отделение и ряд лабораторий,
5
а именно: анатомо-гистологическую, физиологическую, психологическую, биохимическую, большой фотографический кабинет, а позже, в 1908 г., первый в России электрокардиографический кабинет.
Преподавание невропатологии и психиатрии в Военно-медицинской академии и в Женском медицинском институте было поднято В. М. Бехтеревым на большую высоту. Он знакомил студентов со всеми новейшими достижениями в клинике нервных и психических болезней, с перспективами развития этих областей медицины, стремился прививать молодежи материалистическое понимание сущности нервных и психических заболеваний, прогрессивные общественные взгляды.
Научная работа в клинике привлекала множество врачей со всех концов России, нередко из-за границы. За 20 лет руководства клиникой Военно-медицинской академии В. М. Бехтеревым в ней занималось больше тысячи врачей, а ежегодно не менее 40 человек работали над различными научными вопросами в области невропатологии, психиатрии и смежных дисциплин. Под руководством Владимира Михайловича было написано и защищено большое число диссертаций на соискание степени доктора медицины. Исследования, проводившиеся в клинике, докладывались и обсуждались в Обществе психиатров и невропатологов, в Русском обществе нормальной и патологической психологии, на научных заседаниях врачей клиники нервных и душевных болезней Военно-медицинской академии, которыми руководил В. М. Бехтерев, а также в других обществах, на пироговских съездах, на съездах невропатологов и психиатров.
В. М. Бехтерев основал ряд специальных журналов и изданий и являлся их главным редактором. Из них наиболее известны журналы «Обозрение психиатрии, невропатологии и экспериментальной психологии» и «Вестник психологии, криминальной антропологии и гипнотизма». Владимир Михайлович был также членом редакционной коллегии ряда других русских и зарубежных журналов.
За этот период В. М. Бехтеревым были изданы труды «Основы учения о функциях мозга», «Психика и жизнь», «Объективная психология», «Внушение и его роль в общественной жизни» и написано более 200 разнообразных статей, посвященных вопросам анатомии, физиологии, психологии, психиатрии и невропатологии.
Прогрессивные общественно-политические позиции В. М. Бехтерева особенно ярко выявились в период революции 1905 г. Вместе с рядом ученых им был подписан протест, в котором характеризовалось положение школы того времени, освещалась недопустимая политика царизма в области просвещения, опирающаяся на полицейский режим, указывалось на несовместимость академической свободы с государственным строем России, выдвигалось требование политической свободы и народного пред-
6
ставительства в органах государственной власти. В бурный период 1905 г. В. М. Бехтерев по избранию совета Военно-медицинской академии исполнял обязанности начальника академии. В этом же 1905 г. на II съезде отечественных психиатров, происходившем в Киеве, он выступил с докладом «Об условиях развития личности». Этот доклад Бехтерева представлял не только научный интерес —он явился также выражением политического протеста, смысл которого ясно выразился в словах, которыми Бехтерев закончил свой доклад: «Отворите мне темницу, дайте мне сиянье дня». Доклад окончился бурной овацией докладчику, вслед за которой начался митинг, повлекший за собой запрещение съезда.
Широкая известность и авторитет позволили В. М. Бехтереву собрать общественные средства для организации задуманного им нового, прогрессивного научного и учебного учреждения — психоневрологического института и привлечь лучших передовых, демократически настроенных ученых разных областей знания к работе в этом институте.
Задачей основанного в 1908 г. психоневрологического института как научного учреждения было всестороннее изучение личности и разработка вопросов нормальной и патологической неврологии и психологии на широкой основе естественных и общественных наук. Задачей института как высшего учебного заведения являлась подготовка юристов, педагогов и врачей, широко образованных в области естественных и общественных наук и особенно психоневрологии. В институт принимались лица без ограничения пола, возраста и национальности. Студенты через своих представителей в совете института могли участвовать в руководстве жизнью института.
В состав профессоров и преподавателей института входили такие прогрессивные и талантливейшие русские ученые, как Н. Е. Введенский, В. Л. Комаров, П. Ф. Лесгафт, П. А. Останков, Н. Н. Петров, Л. М. Пуссеп, Е. В. Тарле, А. А. Ухтомский и др.
1912 год был годом величайшего гнета со стороны царского правительства и началом нового подъема революционного движения. Речь В. М. Бехтерева, произнесенная в этом году на Московском съезде психиатров, имела важное общественное значение. Говоря о причинах самоубийств, опираясь на официальные данные, он предъявил обвинительный акт самодержавию, показав, что причиной роста самоубийств являются условия жизни царской России, чем вызвал не только широкий сочувственный отклик общественности, но и агрессивную реакцию по отношению к себе со стороны полиции и Министерства просвещения. Воспользовавшись тем, что у В. М. Бехтерева закончился установленный срок службы, его уволили из Военно-медицинской академии; царское правительство не утвердило его президентом созданного и руководимого им института. Наконец,
7
за три дня до февральской революции царское правительство вынесло постановление о закрытии института, которое благодаря революции так и не осуществилось.
Общественная деятельность В. М. Бехтерева выходила далеко за пределы его родины. Он поддерживал активную связь с учеными славянских стран, привлекал их на отечественные съезды, участвовал в организованных ими научных конгрессах. В. М. Бехтерев наметил план связи Российской академии с академиями Балканских стран, разработал единый проект замещения кафедр профессорами в России и других славянских странах, что диктовалось недостатком научных кадров в этих странах и было возможно благодаря близости языков, а главное, разрешало задачи культурного объединения славян и освобождения их от подавляющего немецкого влияния.
Встретив с огромным подъемом Великую Октябрьскую социалистическую революцию, В. М. Бехтерев был одним из первых ученых, которые сразу перешли на сторону молодой Советской республики и отдали все свои силы великому делу построения социализма. Свой доклад на публичном заседании конференции основанного им Государственного института по изучению мозга и психической деятельности в январе 1919 г. В. М. Бехтерев закончил такими словами: «На переломе истории нельзя стоять на перепутье и ждать — нужна воля к действию, к строительству и созидательной работе; и для нас, научных деятелей, которые всегда отдавали свои силы на служение человечеству, не должно быть колебаний. Мы должны отдавать себе отчет, будем ли мы с народом, который, завоевав себе свободу, хочет строить свое будущее сам и зовет нас соучаствовать в этом строительстве. Может ли быть сомнение в ответе на этот вопрос? Мы поэтому должны стремиться к тому, чтобы сократить, по возможности, время разрухи, отдавая всю сумму наших знаний и все умение на созидательную работу в настоящих условиях страны и на пользу народа» г.
1 января 1920 г. было опубликовано в газетах и передано по радио за границу обращение В. М. Бехтерева к врачам всего мира с призывом протестовать печатно и устно против затеянной империалистами Антанты блокады Советской Страны — «неслыханного злодеяния цивилизованных стран, организующих массовое убийство наших граждан, особенно ни в чем не повинных детей и инвалидов» 2.
Огромные возможности для научной, общественной и организаторской работы, которые открылись после Великой Октябрьской социалистической революции, еще больше расширили деятельность В. М. Бехтерева. Совместно с коллективом своих сотрудников он активно включился в разработку вопросов о новой
1 Вопросы изучения и воспитания личности, 1919, № 1, стр. о!.
2 «Петроградская правда» от 1 января 1920 г.
S
культуре, новых задачах здравоохранения, воспитания и т. д.» которые возникли перед молодым Советским государством. Владимир Михайлович руководил созданием новых научных и практических учреждений, организацией научных обществ, съездов, специальных курсов и т. д. В то же время он не оставлял ни лечебной, ни повседневной научной работы, к которой Советское государство предъявляло новые требования. Значительное развитие получил Психоневрологический институт. Молодая советская власть с особым вниманием отнеслась к прогрессивным идеям создателя института и его последователей, направленным на пользу широких трудящихся масс, улучшение здоровья населения, поднятие его культурного уровня на основе широкого развития советской науки.
Научная деятельность В. М. Бехтерева так обширна и многостороння, что не представляется возможным перечислить хотя бы названия его научных работ, а тем более достаточно полно осветить их значение.
Исследования В. М. Бехтерева по анатомии, напечатанные в различных русских и иностранных изданиях, были объединены в большом труде «Проводящие пути спинного и головного мозга». За этот труд он получил премию Бера Российской академии наук. В этой работе В. М. Бехтерев впервые дал полное и систематическое описание связей, соединяющих различные отделы спинного мозга, и хода нервных путей в различных отделах центральной нервной системы, представил систему строения головного и спинного мозга. Он показал неодновременность развития и тем самым разную функциональную роль различных проводящих систем и неодновременность созревания различных клеточных образований мозга, из которых корковые клетки развиваются позже всех.
В. М. Бехтерев убедительно опроверг положение Вирхова: «каждая клетка из клетки», доказав, что кора головного мозга в раннем периоде не имеет клеточного строения х. В этом отношении его работы близки к современным исследованиям О. Б. Ле-пешинской.
В. М. Бехтереву также принадлежит ряд плодотворных идей и исследований в области динамической морфологии. Так, им совместно с сотрудниками были произведены исследования изменения структуры мозговых клеток в связи с изменением их деятельности (В. И. Нарбут), указана изменчивость шиловидных отростков (синапсов), осуществляющих контакт между нев-ронами.
Начатое под руководством В. М. Бехтерева еще в Военнс-медицинской академии изучение окончаний нервов во внутрен-
1 Обозрение психиатрии, невропатологии и экспериментальной психологии, 1899, № 2.
9
них органах (С. А. Михайлов) было затем продолжено и развито в ряде работ отдела морфологии руководимого В. М. Бехтеревым Института мозга и вылилось в обширный круг вопросов о связи внутренних органов с центральной нервной системой.
В процессе морфологических исследований В. М. Бехтеревым открыт ряд нервных образований; некоторым из них присвоено имя В. М. Бехтерева. Так, им были описаны: ход задних корешков и их расчленение на отдельные группы; названная псзже его именем группа клеток в наружной части шейки заднего рога; три ранее неизвестных пучка в спинном мозгу. В. М. Бехтерев подробно изучил так называемое сетевидное образование, описал ряд нервных пучков и ядер в мозговом стволе. Одно из них — ядро вестибулярного нерва — носит его имя. Изучая строение мозжечка, В. М.. Бехтерев установил связи червячка и центральных ядер мозжечка как рецепторов центростремительных импульсов, исходящих с периферии (суставы, мышцы и сухожилия, полукружные каналы); он показал, что полушария мозжечка связаны с варолиевым мостом и большими полушариями. Он же описал четыре пучка в brachia conjunctiva и установил, что striae medullares служат для соединения различных частей мозжечка.
В. М. Бехтерев показал, что бледное ядро соединяется с мозговой корой большими пучками, в то время как хвостатое тело и скорлупа получают из коры коллатеральные пучки, проходящие, вероятно, через внутреннюю капсулу. Он показал также, что наружный отдел мозговой ножки, считавшийся раньше центростремительным, на самом деле является центробежным проводником. Наружный корешок обонятельного нерва идет, по Бехтереву, прямо в крючковидную извилину, бахрома, а также свод являются центрифугальными пучками. В наружных слоях мозговой коры В. М. Бехтеревым найдена полоска, которая названа его именем.
Заслуживает большого внимания открытая им особенность строения мозговой коры, заключающаяся в том, что во всех участках ее имеются двоякого рода проводники — нисходящие и восходящие.
В. М. Бехтереву принадлежит ряд работ по физиологии, в которых теснейшим образом сочетаются вопросы морфологии и физиологии, клиники и экспериментального исследования. Так, совместно с П. Я. Розенбахом он установил трофические функции спинномозговых узлов.
Важное место занимают исследования В. М. Бехтерева в об" ласти функции органов равновесия. На основании экспериментов, имеющих не только физиологическое, но и клиническое значение, установлена роль полукружных каналов и лабиринта, дна третьего желудочка и мозжечка.
Большое значение имеет открытие В. М. Бехтеревым функции зрительных бугров, их роли в деятельности внутренних органов и в выразительных реакциях. Значение зрительных бугров как
10
подкоркового вегетативного центра освещено им задолго до работ Карплуса и Крейдля.
Нельзя не отметить роли В. М. Бехтерева в развитии электроэнцефалографии. Работы, вышедшие из его лаборатории (В. Е. Ларионов, С. А. Тривус, П. Ю. Кауфман), показали, что биоэлектрические явления обнаруживаются не только в коре полушарий, но что они не одинаково выражены в различных областях мозга в зависимости от характера воздействий и деятельности мозга.
В ряде работ совместно с проф. Н. А. Миславским и самостоятельных, а также на основе исследования ряда учеников В. М. Бехтерев установил влияние раздражения предлежащих участков коры головного мозга на деятельность многих внутренних органов. Физиологические исследования В. М. Бехтерева подытожены в его большой работе.состоящей из 7 томов: «Основы учения о функциях мозга», представляющей энциклопедию экспериментальной и клинической физиологии мозга на этапе методов экстирпации и раздражения. В своем отзыве об этой работе И. П. Павлов указывал, что «она представляет изложение, единственное по своей полноте не только в русской, но и в иностранной литературе», основанное не только на литературном знании, но и на личном опыте. Считая что эта книга «должна стать главным руководством при специальном ознакомлении врачей и естественников с центральной нервной системой», И. П. Павлов делает ряд важных критических замечаний, из которых особенно существенны замечания, относящиеся к деятельности высших отделов центральной нервной системы, указывающие на смешение В. М. Бехтеревым психологической и физиологической точки зрения, на встретившее в литературе оппозицию представление о корковых центрах и на недостаточность методики их исследования. Применявшаяся В. М. Бехтеревым методика не давала возможности установить физиологические механизмы связи коры головного мозга и внутренних органов.
А. Г. Иванов-Смоленский об этом руководстве пишет: «Здесь дана находящаяся на уровне знаний того времени, почти исчерпывающая сводка исследований, посвященных локализации рецепторно-сензорных, вегетативных, скелетно-двигательных, сочетательных и высших психических функций в различных отделах мозга» х. В это время И. П. Павлов уже создал свой классический метод условных рефлексов, обеспечивающий строго объективное исследование высшей нервной деятельности животного и позволивший ему открыть корковую условнорефлектор-ную регуляцию деятельности внутренних органов и тканевых процессов. Этот вопрос впоследствии был широко разработан рядом его сотрудников, особенно М. К. Петровой, К. М. Быковым и М. А. Усиевичем.
1 Журнал высшей нервной деятельности, 1953, т. II, в. 6.
11
В. М. Бехтерев в дальнейшем основывается на методе и учении об условных рефлексах, называя их сочетательными. Он писал: «Неприменимость (по тому времени.—В. М.) слюноотделительной методики к человеку заставили нас искать возможность выработать для лабораторных исследований сочетательно-двигательный рефлекс» х.
Нет необходимости говорить о том, что термин «сочетательный рефлекс» представляет синоним условного рефлекса, хотя термин «условный рефлекс» правильнее отражает лежащую в основе его временную связь между очагами возбуждения, возникающими в коре головного мозга при действии раздражителей на организм, тогда как термин «сочетательный рефлекс» не раскрывает сущности высшей нервной деятельности.
Первая публикация об исследовании условных двигательных рефлексов у человека на основе исследования глотательного рефлекса у детей принадлежит одному из виднейших учеников И. П. Павлова — Н. И. Красногорскому 2. В. М. Бехтерев с сотрудниками сначала сделал попытку образования условных дыхательных рефлексов как у собак, так и у людей (В. Я. Анфи-мов, И. П. Спиртов), а затем остановился на методике двигательного условного рефлекса на животных и на людях (В. П. Протопопов, Н. И. Добротворская, А. Г. Молотков и др.).
В дальнейшем В. М. Бехтерев трактует все материалы своего прошлого опыта, клинических проблем и психической деятельности человека, основываясь на рефлекторной теории и прежде всего на учении об условных рефлексах.
В физиологических исследованиях на животных В. М. Бехтерев с самого начала, исходя из интересов неврологической клиники, был занят вопросами локализации, разрешая их с помощью методов раздражения и экстирпации.
Но в дальнейшем в понимании как физиологических, так и клинических фактов он все более преодолевает статические односторонне морфологические взгляды современных ему невропатологов и приближается к динамическому физиологическому пониманию, которое было последовательно развито в физиологическом учении И. П. Павлова о высшей нервной деятельности.
Так, установленную его исследованиями роль коры головного мозга в деятельности внутренних органов он начинает трактовать как условнорефлекторную (сочетательнорефлекторную) 3. Корковые центры В. М. Бехтерев первоначально рассматривал как место проекции афферентных и исходное место эфферентных
1 В. М. Бехтерев, Русский врач, 1909, № 31, 33 и 36.
2 Н. И. Красногорский, Русский врач, 1907, № 36.
3 В. М. Бехтерев, Исследование функций мозговой коры с помощью естественных сочетательных рефлексов. Значение этого метода по-отношению к центрам внутренних органов и различных секреций, Обозрение психиатрии, невропатологии и экспериментальной психологии, 1908, № 8.
12
импульсов. Затем он выдвинул понимание их как «диференциру-ющих площадок» в коре головного мозга. Размеры их различны для различных органов, а главное, динамически изменяются в зависимости от степени возбудимости. В. М. Бехтерев указывал на то, что центр представляет сложное образование, расположенное в различных этажах нервной системы. Если раньше он рассматривал центры только как место проекции того или иного органа, то в дальнейшем говорит о них как о местах образования связей между раздражителем и ответной реакцией. При этом Бехтерев различает первичные центры, где образуются простые сочетания раздражений одного рецептора, и вторичные, в которых возникают сочетания сложных разнородных раздражений. Ассоциационных центров Флексига Бехтерев не признавал, так же как и И. П. Павлов.
В настоящее время работы В. М. Бехтерева, основанные преимущественно на методах раздражения и экстирпации, имеют главным образом историческое значение. Однако собранный в этих исследованиях большой фактический материал, объединяющий анатомию, физиологию и клинику, до сих пор имеет ценность для разрешения важнейшей проблемы связи структуры к функции, значение которой неоднократно подчеркивалось И. П. Павловым и разрешение которой сейчас осуществляется на основе созданной И. П. Павловым физиологии головного мозга.
Работы В. М. Бехтерева по психологии вначале не выходили за рамки объективного экспериментального психо-физиоло-гического исследования и были направлены преимущественно на экспериментальное изучение вопросов чувствительности, восприятия, ассоциативных процессов, моторных актов. Лишь с конца 90-х годов В. М. Бехтерев приступил к перестройке системы психологии на основе объективного метода. Основным его трудом в этой области является «Объективная психология», в которой делается попытка построить систему объективно-материалистического учения о рефлексах, особенно тех, которые приобретаются в индивидуальном опыте, т. е. условных или, как он называл их, сочетательных рефлексов.В дальнейшем В. М. Бехтерев заменил термин «объективная психология» термином «психорефлексология» и позднее ввел термин «рефлексология». Различные виды психической или высшей нервной деятельности В. М. Бехтерев рассматривал как различные виды условных (сочетательных) рефлексов.
Бесспорную заслугу В. М. Бехтерева представляет его стремление найти за мистифицированной системой понятий субъективно-идеалистической психологии их объективное содержание, заключающееся в актах взаимодействия индивидуума со всей окружающей его действительностью.
Однако, стоя в основном на материалистической позиции объективного исследования, В. М. Бехтерев не был вооружен
13
единственно правильной методологией марксизма-ленинизма, поэтому он полностью не преодолел идеалистических и механистических ошибок, не был достаточно последователен в своих материалистических позициях, обнаруживал эклектизм. Борясь с вульгарным материализмом, он не был знаком с диалектическим материализмом; критикуя энергетизм Оствальда, Лас-свица и других, он свои взгляды называл также энергетизмом.
В. М. Бехтерева не удовлетворяла старая субъективная психология. Разрабатывая объективную психологию, он в вопросе о взаимоотношении между психическими и физиологическими процессами высказал (в своей работе «Психика и жизнь», 1902) «энергетическую концепцию», ничего общего не имевшую с диалектическим материализмом.
Стоя на позициях объективно-материалистического исследования личности, В. М. Бехтерев допускал возможность существования, наряду с объективной психологией, которую он позднее назвал рефлексологией, субъективной психологии, т. е., даже борясь с параллелизмом, он не мог полностью преодолеть его и стать на позиции последовательного материалистического монизма.
О недостаточности естественно-научного материализма В. М. Бехтерева свидетельствуют наиболее серьезные его ошибки в объяснении общественных явлений как рефлекторных актов, допущенные им в работе «Коллективная рефлексология».
Рефлексологическая концепция В. М. Бехтерева резко отличалась от созданной И. П. Павловым физиологии высшей нервной деятельности. Она возникла в результате попытки эклектически использовать физиологию головного мозга и данные субъективной психологии. Противопоставление явлений «нервного тока», т. е. «сочетательных» рефлексов, субъективным психологическим процессам привело В. М. Бехтерева к высказываниям, относящимся к психо-физиологичес-кому параллелизму. На ложных методологических позициях была создана Бехтеревым и так называемая коллективная рефлексология.
В работе «Коллективная рефлексология» В. М. Бехтерев пытался установить общие для мертвой материи, органической природы и общества законы, например, законы инерции, тяготения и др. Несмотря на признание их качественного различия, попытка распространить одни и те же законы и на явления неорганического и животного мира, и на общественные явления свидетельствует о непонимании Бехтеревым своеобразия этих явлений, о механицизме, биологизме и психологизме, особенно ясно выступающих при разрешении им социальных проблем.
Оттеняя преимущественную роль в развитии человека общественных условий, В. М. Бехтерев говорил о биосоциальной природе человека. При этом он не учитывал, что такое определение приемлемо и для эклектической теории двух факторов, не пытался
14
отмежеваться от нее и стать на позиции последовательного исто-рико-материалистического понимания общественной природы человека.
Будучи выдающимся клиницистом невропатологом и психиатром, В. М. Бехтерев выдвигал важные прогрессивные и чрезвычайно ценные и для настоящего времени общие медицинские идеи.
То направление, которого придерживался В. М. Бехтерев в медицине, нередко называют анатомо-физиологическим. Однако такая характеристика его взглядов не может быть признана вполне правильной. Об этом свидетельствует ряд высказываний В. М. Бехтерева, в которых он указывает, что «следует совершенно отказаться от анатомического принципа объяснения психических расстройств...» и что «основа психических болезней должна быть патологофизиологическая, а не патологоанатоми-ческая...» *.
Критикуя целлюлярную патологию Вирхова, В. М. Бехтерев указывал на то, что психические болезни, выражающие «нарушения "отправлений мозга, лишь относительно редко стоят в связи с анатомическими нарушениями самого мозга» 2. «Центр тяжести болезненного процесса в огромном большинстве душевных заболеваний лежит вовсе не в изменениях мозга» (имеются в виду анатомические изменения.—В. М.), и даже по отношению к заболеваниям с «более или менее очевидными, сравнительно легко открываемыми патологоанатомическими изменениями мозга мы не видим в самом мозгу корней заболевания, а должны искать их в нарушенных условиях жизнедеятельности всего организма»3. Таким образом, в основу психоза кладется единство нарушенной деятельности мозга и всего организма, единство соматического и психического. В единстве организма, помимо регулирующей его жизнедеятельность центральной нервной системы, в частности, коры головного мозга, В. М. Бехтерев подчеркивал значение «биохимических систем». Эти биохимические системы связаны главным образом с эндокринными железами, роль которых и связь их с нервной системой были оценены В. М. Бехтеревым на заре эндокринологии. Также понимал он и значение обмена веществ.
В. М. Бехтерев указывал на то, что « в происхождении приобретаемых под влиянием внешних условий особенностей организма играет роль тот же гормонизм, который лежит в основе прирожденных отличительных признаков». При внешних воздействиях функциональная роль тех или иных железистых органов зависит от «внешних условий развития при известных данных питания, температуры, солнечной энергии, большей или мень-
1 Обозрение психиатрии, невропатологии и экспериментальной психологии, 1908, № 1.
2 Там же.
3 Русский врач, 1912, № 6.
15
шей влажности окружающего воздуха и т. п.» х. Соответственно этому В. М. Бехтерев решительно утверждал в согласии с И. М. Сеченовым и И. П. Павловым наследственное закрепление приобретенных особенностей.
Исходя из рефлекторной теории, В. М. Бехтерев освещал генез заболеваний, включающий как более элементарные, так и сложнейшие рефлекторные механизмы психических заболеваний. Основываясь на учении об условных (сочетательных) рефлексах, В. М. Бехтерев освещал патогенез психозов, их диагностику и терапию.
Заслуги его во внедрении теории условных рефлексов в клинику справедливо отметил А. Г. Иванов-Смоленский: «Впервые учение о высшей нервной деятельности было распространено на человека в виде начальных попыток экспериментального исследования корковой динамики у детей в 1907 г. Н. И. Красногорским (одним из старейших учеников И. П. Павлова). Но первый опыт введения этого учения в область невропатологии и психиатрии принадлежит (1908—1909) В. М. Бехтереву и его ученикам. Можно, и даже должно, не соглашаться с ним в его склонности, с одной стороны, к чрезмерному упрощению столь сложной задачи, а с другой стороны, к слишком широким и неправомерным обобщениям при попытках разрешения ее, но вместе с тем необходимо признать следующее: непререкаемая и бесспорная заслуга В. М. Бехтерева состояла в том, что он был первым представителем невропатологии и психиатрии, правильно оценившим огромные перспективы, открываемые учением И. П. Павлова для этих медицинских дисциплин. В этом его несомненная и большая заслуга перед отечественной наукой4» 2.
В. М. Бехтерев является также крупнейшим представителем общественно-гигиенического направления в психиатрии и невропатологии.
На заре своей деятельности он обнаружил правильное понимание значения общественных условий для здоровья, возникновения болезни и борьбы с ней и с большой смелостью высказывал эти взгляды при царском режиме. Уже в 1885 г. в Казани в своем первом публичном выступлении он указывал на тяжелые экономические условия жизни населения в царской России как на причину распространения психических заболеваний. Он правильно утверждал, что в основе нервно-психической заболеваемости населения лежит «экономическое рабство, обусловленное значением капитала в жизни современного общества. Капиталистический строй — вот основное зло нашего времени»2.
1 Русский врач, 1913, № 7.
2 А Г. Иванов-Смоленский, Очерки [патофизиологии высшей нервной деятельности, Медгиз, 1952, стр. 258.
3 Вопросы невро-психического здоровья в населении России, 1911.
16
Мысль о пагубных условиях капиталистического строя высказывалась В. М. Бехтеревым неоднократно. Так, в 1908 г. он говорил, что все усилия в борьбе с вырождением населения и за улучшение жизненного уровня населения «должны быть направлены на устранение капиталистического режима»1. В 1912 г. он пишет:«..несама культура и цивилизация,не умственная работа и просвещение масс, не мозговое переутомление служат условиями, приводящими в современном обществе к развитию душевных болезней, а те отрицательные стороны, которые ослабляют организм вообще и связаны с капиталистическим строем общества»2.
Эти слова В. М. Бехтерева, произнесенные им на больших собраниях в период господства царской реакции, когда революционное движение было загнано в подполье, свидетельствуют о глубине его мысли, о мужестве, с каким он отстаивал свои взгляды и интересы народных масс.
Принципиальные клинические позиции В. М. Бехтерева складывались в процессе разносторонних конкретных исследований. Здесь В. М. Бехтереву принадлежат многочисленные открытия, принесшие ему заслуженную славу. Остановимся лишь на наиболее важных из них.
В области невропатологии В. М. Бехтеревым описаны нормальные и патологические рефлексы: бехтеревский (сгибательно-пальцевой), ранее неправильно называвшийся мендель-бехте-ревским, лопаточный, метакарпальный, тибиальный, сакролюм-бальный, орбикулярный, носовой, ментальный, губной, щечный, гипогастрический или субингвинальный. Им описан ряд симптомов ранней стадии сухотки спинного мозга, заболеваний основания мозга, невралгии седалищного нерва, невритов. Вместе с П. А. Останковым им был описан симптом Pseudomelia para-esthetyca, вместе с Г. Е. Шумковым — симптом мелких шагов при паркинсонизме.
К впервые описанным В. М. Бехтеревым болезненным формам в клинике нервных болезней относятся: бехтеревская форма неподвижности позвоночника, сифилитический множественный склероз, апоплектическая гемитония, хореическая падучая, акроэритроз, особая форма лицевого тика, острая мозжечковая атаксия при алкоголизме, невроз местной контузии и др.
В психиатрической клинике им выделено значительное число синдромов: бред «гипнотического очарования»; ряд форм с галюцинациями периферического происхождения (вследствие поражения шнейдеровской мембраны, болезней среднего уха, поражения слизистой оболочки носа и т. д.); рептилофрения (одержимость гадами), психические нарушения в связи с раздражением со стороны желудочно-кишечного тракта; сомато-френия и др.
1 Вопросы вырождения и борьба с ним, 1908.
2 Основные задачи психиатрии как объективной науки, Русский врач, 1912.
2 В. М. Бехтерев
17
Большая роль принадлежит В. М. Бехтереву в разработке вопроса о неврозах и пограничных состояниях. Так, им впервые описаны объективные признаки расстройств чувствительности при неврозах на основе изменения вегетативных, в частности, сосудистых, потовых, температурных рефлексов в областях измененной чувствительности (1893). Им же предложена на основе условных рефлексов диагностика симуляции, функциональных и органических параличей и расстройств чувствительности. В. М. Бехтереву принадлежит описание разнообразных обсес-сивных нарушений. Он же предложил применить учение об условных рефлексах для объяснения природы обсессий, половых извращений и ряда истерических расстройств. Заслуживает особого внимания то, что В. М. Бехтерев для объяснения истерических расстройств привлек теорию парабиоза Н. Е. Введенского. Различные состояния парабиоза соответствуют переходным фазам или гипноидным фазам И. П. Павлова.
В. М. Бехтерев широко применял принцип условных рефлексов при лечении ряда болезненных состояний. Сюда в первую очередь должны быть отнесены лечение расстройств чувствительности и движения при неврозах, а также лечение наркомании, особенно алкоголизма и курения. Как известно, еще в 1915 г. В. М. Бехтерев предложил лечение алкоголизма путем сочетания алкоголя с рвотным средством *.
Благодаря открытиям В. М. Бехтерева отечественная психоневрология заняла одно из ведущих мест в мировой науке. Однако ряд открытий В. М. Бехтерева до сих пор не связывается с его именем и часто приписывается зарубежным авторам.
Можно, например, указать, что агнозия пальцев, которая считается установленной Герстманом, описана Бехтеревым в его труде «Основы учения о функциях мозга» 2; синдром психического автоматизма, приписываемый Клерамбо, правильнее называть именем Кандинского-Бехтерева 3; описание подкорковой эпилепсии принадлежит не Пенфильду, а В. М. Бехтереву и Иценко 4; синдром поражения боковой нижней части продолговатого мозга описан впервые Бехтеревым, и лишь через 10 лет он был описан Валленбергом и неправильно получил его имя5; сгибательный рефлекс ноги, описанный Бехтеревым в 1900 г., считается «открытым» Мари и Фуа 6 в 1910 г.
В своей работе о роли органов равновесия в восприятии пространства В. М. Бехтерев явно предвосхищает идеи Магнуса о
1 Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915, № 10—12.
2 Основы учения о функциях мозга, 1906, т. VI, стр. 826.
3 Обозрение психиатрии, невропатологии и экспериментальной психологии, 1896, № 10.
4 Там же.
5 Нервные болезни в отдельных наблюдениях, 1894.
β Обозрение психиатрии, невропатологии и экспериментальной психологии, 1900, № 9.
18
рефлексах положения 1. Современные энцефало- и ангиография имеют свои корни в заявлении В. М. Бехтерева о том, что можно будет изучать состояние мозга и его сосудов, вводя в сосуды вещества, не пропускающие рентгеновых лучей2.
В. М. Бехтерев является крупнейшим практиком-психиатром. Он посвятил многочисленные работы и осуществил ряд мероприятий по организации системы нестеснения и улучшения режима психиатрических учреждений, внедрению в психиатрическую практику многообразных видов терапии, организации культ-и трудотерапии в психиатрических учреждениях, разработке вопросов профилизации психиатрических отделений, призрения и патронажа психически больных, организации психо-невроло-гических диспансеров. В этой деятельности В. М. Бехтерева тесно связывались лечебно-организационная практика и гуманные ■ общественные идеи.
В терапии психических болезней В. М. Бехтереву свойственна большая активность и огромная настойчивость, с которой он боролся за восстановление здоровья нервно- и психически больных. Он применял все, что давала ему современная наука. Физиотерапия, механотерапия и психотерапия, рентгенотерапия и нейрохирургия, фармакотерапия, трудотерапия, серотерапия, опотерапия и др.—весь этот богатый арсенал средств использовался гибко и диференцированно с соблюдением принципа гуманности и осторожности в отношении к больному. В. М. Бехтерев был одним из активнейших организаторов рентгенотерапии нервно- и психически больных, цвето- и светолечения. Большое внимание он уделял лечению музыкой — метод, до сих пор недостаточно развитый и оцененный.
В фармакотерапии В. М. Бехтерев проводил принцип комбинированного действия рядом лекарственных веществ. Примером такой комбинации является его знаменитая микстура. Он активно внедрял трудовую терапию. Творческая инициатива В. М. Бехтерева в области психотерапии проявлялась во введении ряда методических вариантов психотерапии — гипноза, лечения отвлечением.
В. М. Бехтерев сделал чрезвычайно много для пропаганды и популяризации науки. Он опубликовал многочисленные популярные статьи по вопросам воспитания, гигиены нервной системы, научного понимания нервно-психической деятельности.
Будучи выдающимся организатором, В. М. Бехтерев привлекал к себе широкий коллектив, на который он всегда опирался в своей работе. Из коллектива сотрудников В. М. Бехтерева вышли выдающиеся советские невропатологи, психиатры и психологи.
1 Вестник клинической и судебной психиатрии, 1884, № 1.
2 Отчеты научных собраний Санкт-Петербургской клиники нервных и душевных болезней за 1895—1896 и 1896—1897 гг.
2*
19
В кипучей деятельности В. М. Бехтерева сочетались теория с практикой, изучение с лечением и воспитанием. Ученый-общественник, он объединял в себе индивидуальные и общественные устремления, личное творчество с трудом на благо народа.
Стоя на рубеже двух эпох, он соединял в себе лучшие традиции русской дореволюционной научной интеллигенции с энтузиазмом молодого социалистического поколения. Он первый из крупнейших русских ученых после статьи В. И. Ленина «Великий почин» откликается на нее статьей, в которой с подъемом пишет об огромном значении субботников, видя в них проявление творчества народных масс, «шествие труда свободного, коллективного, бескорыстного, почти героического».
Когда в Америке разразился знаменитый своим обскурантизмом «обезьяний процесс», В. М. Бехтерев немедленно откликается на него статьей «Борьба за науку». Бичуя фанатиков церкви и политически реакционные группы американских шовинистов, он заявляет: «Материалистические выводы и положения современной науки должны быть самым решительным образом выдвинуты на борьбу с грозной эпидемией фанатического изуверства, идущей с дальнего запада из Америки, кичащейся своими техническими достижениями и свободами» г. Как близки эти слова нашей современности и о какой чуткой прозорливости они свидетельствуют !
В. М. Бехтерев является выдающимся деятелем отечественной науки и культуры, тружеником, которым всегда будет гордиться наша великая социалистическая родина.
Трудящиеся Ленинграда оказали В. М. Бехтереву высокое доверие, избирая его при каждых выборах депутатом Петроградского, в дальнейшем Ленинградского совета депутатов трудящихся.
Умер В. М. Бехтерев 24 декабря 1927 г. в полном расцвете творческих сил.
В речи, произнесенной у гроба В. М. Бехтерева, председатель ЦИК М. И. Калинин дал высокую оценку его деятельности. Товарищ Калинин сказал: «Осознание единства науки и социализма и практическое претворение в жизнь этого сознания — вот что является одной из огромнейших заслуг покойного. Академик Бехтерев много помог сближению труда и науки и этим самым укреплению рабоче-крестьянского строя» 2.
В. Мя с ище в
1 Железнодорожная техника и экономика, 1919, № 3. • «Известия» от 25 декабря 1927 г.
20
I
ТРУДЫ ПО АНАТОМИИ МОЗГА
ЭКСПЕРИМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ
ОТНОСИТЕЛЬНО ПЕРЕКРЕСТА ВОЛОКОН
ЗРИТЕЛЬНОГО НЕРВА В CHIASMA NN. OPTICORUM
Хотя уже давно на основании некоторых фактов клинической патологии было высказано предположение о неполном перекресте зрительных нервов в chiasma человека, тем не менее, несмотря на обширный казуистический материал, накопившийся со времени Уоллестона, упомянутый вопрос до сих пор еще может считаться спорным, и лица, держащиеся мнения о неполном перекресте зрительных волокон, принимают его не иначе, как за гипотезу, при помощи которой становится удобным объяснять существование половинной односторонней слепоты при мозговых поражениях, помещающихся в области, соседней с tractus optici. Но и между клиницистами до сих пор еще не существует полного согласия по этому вопросу. Некоторые из новейших авторов, как Дикинсон 1, вопреки многим, высказываются еще за существование полного перекреста.
Такое противоречие между авторами по вопросу, всегда занимавшему видное место в медицинской литературе, показывает с ясностью, в какой степени могут быть шатки воззрения, выведенные на основании одних фактов клинической патологии, и С какой осторожностью и осмотрительностью следует пользоваться клиническим материалом. Правда, в решении указанной задачи были принимаемы в соображение также и анатомические данные, но, к сожалению, и здесь результаты многочисленных исследований далеко не привели к согласным выводам. Прочно установленным может считаться только то, что у низших животных, как рыбы, земноводные и птицы, существует полный перекрест зрительных волокон в chiasma. Что же касается высших животных и, между прочим, человека, то анатомические исследования различных авторов привели к совершенно противоречивым данным. В то время как Ганновер 2, Генле 3 и в последнее
1 Dickinson, Optic-chiasm. Visual centres, The alienist and neurologist, 1881, p. 358.
2 Hannover, Das Auge, Beitrage zur Anatomie, Physiologie u. Pathologie dieses Organs, Leipzig, 1872.
8 H e η 1 e, Nervenlehre. Ueber die Kreuzung in chiasma ηη. opticorum.
23
время Гудден1 считали возможным установить существование неполного перекреста, другие авторы, как Бизиадецкий2>Мандель-штам3 и Мишель4, высказались в противоположном смысле, т. е. за существование полного перекреста.
С целью примирить эти разноречивые взгляды некоторые авторы в отношении перекреста волокон в chiasma предполагали даже существование индивидуальных различий, подобно тому как существуют количественные различия перекреста волокон в пирамидах (Флексиг). Подобное предположение в отношении chiasma, впрочем, до сих пор еще не имеет за себя никаких положительных данных.
Из числа анатомических работ исследования Гуддена, несомненно, заслуживают наибольшего внимания ввиду того, что они были выполнены по принципу экспериментальной гистологии. Автор, разрушая bulbi oculi новорожденным животным, мог выделить из всего ствола tractus opticus те волокна, которые подвергались последовательному перерождению, от тех, которые избегли подобного перерождения. На основании полученных результатов Гудден мог констатировать, что каждый tractus opticus состоит из четырех частей: 1) волокон, которые, собственно говоря, не принадлежат самому tractus, а только сопровождают его на некотором протяжении, входя в состав так называемой спайки Мейнерта; 2) волокон, которые принадлежат самому tractus, но не входят в состав п. optici, а, переходя в заднем углу chiasma с одной стороны на другую, составляют так называемую commissuram infer. Guddeni; 3) волокон, переходящих из tractus через chiasma без перекреста, которые лежат в наружной части соответствующего п. optici, и 4) волокон, перекрещивающихся в chiasma и занимающих самую внутреннюю часть п. optici.
Эти исследования, таким образом, говорят в пользу существования неполного перекреста волокон п. optici в chiasma более высших позвоночных, хотя они не дают еще права утверж? дать, что количественное отношение перекрещивающихся и не перекрещивающихся волокон у высших животных должно быть то же самое, как и у человека.
Несмотря на совершенство приведенных исследований Гуддена, многие авторы признают вполне справедливо, что анатомические данные не могут еще служить к окончательному выяснению спорного вопроса, решение которого всецело находится в руках экспериментальной физиологии.
1 Gudden, Arch. f. Ophtalmologie, 1874, Bd. 25.
2 Biesiadecki, Ueber das Chiasma nervorum opticorum d. Men-sc.hen u. d. Thiere, Wiener Sitzungsb. d. Mathem. u. Naturwiss., 1861, Bd. XLII.
•Mandelstamra, Arch, f. Ophtalm., 1873, Bd. XVI. 4 M i s с h e 1, Ibid.
24
Действительно, вопрос о перекресте зрительных волокон в chiasma мог быть решен экспериментальным путем весьма просто, если бы удалось сделать перерезку или одного из tractus opticus, или разрезать всю chiasma ηπ. opticorum в передне-заднем направлении. Такие исследования производились, но, к сожалению, и они не привели к согласному результату. Кнолль г первый начал исследования подобного рода. Производя в одном ряде опытов перерезку п. optici и убедившись, что у животного под влиянием указанной операции наступало вместе со слепотой соответствующего глаза расширение зрачка и неподвижность его при непосредственном раздражении светом, автор произвел в другом ряде опытов перерезку tractus opticus, причем наблюдал те же явления, но только на противоположном глазу. Несколько позже Броун-Секар 2 опубликовал результаты своих опытов над кроликами и морскими свинками, которым он производил как перерезку одного из tractus opticus, так и рассечение chiasma в передне-заднем направлении, а также разрушение согр. genicul. и согр. quadrigemini. Во всех этих случаях автор наблюдал различного рода нарушения зрительной способности: то полную слепоту соответствующего или противоположного глаза или обоих вместе, то половинную слепоту одного или одностороннюю половинную слепоту обоих глаз. При этом он не определяет в подробностях, каким поражениям соответствует то или другое нарушение зрения, но утверждает, что перерезка chiasma в передне-заднем направлении вызывала полную слепоту обоих глаз, а перерезка одного tractus — полную слепоту противоположного глаза 3.
Оба приведенные исследования говорят, таким образом, в пользу существования полного перекреста зрительных волокон в chiasma ηη. opticorum. Напротив того, исследования Никати* привели к совершенно противоположному результату. Производя рассечение chiasma в передне-заднем направлении у кошки, автор не наблюдал прекращения зрительной способности как со
1 Knoll, Beitr. z. Anat. u. Physiol, v. Eckhard, Giessen, 1869, Bd. IV.
• Brown-Sequard, Arch, de physiol. norm, et pathol., 1872, vol. IV, 2, p. 261.
8 Автор, впрочем, придерживается совершенно особенного взгляда относительно происхождения упомянутых зрительных расстройств. По его мнению: П одного полушария мозга достаточно для зрения обоими глазами, и каждый tractus opticus, будучи связан с соответствующим полушарием мозга, находится в сообщении с обеими половинами обеих сетчаток; 2) amaurosis в случае повреждения tractus opticus, corp. quadrig. или других частей соответствующего мозгового полушария зависит не от прекращения функции проводников зрительных впечатлений или центра, воспринимающего эти впечатления, но от влияния на питание глаза или п. optici, вследствие раздражения, происходящего в месте повреждения. Подобный взгляд, сколько мне известно, до сих пор, однако, не разделяется никем из современных физиологов и в пользу его не существует пока никаких положительных фактов.
* Nicati, Zbl. f. med. Wissensch., 1878, S. 449.
25
стороны того, так и со стороны другого глаза, из чего можно заключить только о существовании у оперированных животных неполного перекреста волокон в chiasma ηη. opticorum.
Недостаток приведенных исследований очевиден уже из того, что экспериментаторы производили свои опыты над такого рода животными, относительно которых уже анатомические данные заставляют сомневаться в существовании неполного перекреста зрительных волокон в chiasma. По моему мнению, вопрос этот может быть решен окончательно только на основании экспериментальных исследований, произведенных над более высшими животными, как, например, собаками, над которыми подобных исследований еще не производилось.
Способ, при помощи которого удается достигнуть рассечения tractus opticus у такого рода животных, может быть двояким: один состоит в том, что проникают обоюдоострым ножом из пасти животного внутрь черепа в месте, соответствующем турецкому седлу, и перерезывают или самый tractus opticus, или, что удается только в некоторых счастливых опытах, рассекают chiasma пп. opticorum в передне-заднем направлении. Другой способ требует трепанации черепа тотчас над arcus zygomaticus, в промежутке между глазом и ухом животного, после чего внутрь черепа проводят загнутый по краю нож с притуплённым концом и, скользя им по внутренней поверхности средней черепной ямки с целью обойти височную область мозгового полушария, достигают, наконец, боковой стенки турецкого седла; на месте последней поворачивают нож лезвием кверху и производят перерезку tractus opticus одним движением ножа в обратном направлении; затем снова поворачивают нож плашмя и, держась стенки черепной покрышки, осторожно выводят его обратно.
Каждый из этих способов имеет свои выгоды и невыгоды. Первый, несомненно, имеет преимущество в том отношении, что дает возможность произвести вполне изолированную перерезку не только tractus opticus, но и opticus (внутри черепной полости), а также сделать рассечение chiasma в том или другом направлении. Невыгода же этого способа состоит в том, что он требует особенного навыка и при всем том, пользуясь этим способом, не всегда удается действовать наверняка. При употреблении второго способа, напротив того, почти всегда можно рассчитывать на успех в отношении перерезки tractus, но при этом обыкновенно в большей или меньшей степени повреждается также и соседняя ■мозговая ткань. Впрочем, от руки экспериментатора зависит свести эти посторонние повреждения до возможного минимума.
Результаты сделанных мною опытов над собаками по тому и другому способу заключались в следующем.
При перерезке п. optici у животного вместе с полной слепотой соответствующего глаза наступало значительное расширение зрачка и неподвижность его по отношению к свету, если последний падает непосредственно на исследуемый глаз. Напротив
26
того, если свет проникает через здоровый глаз, то зрачок глаза с перерезанным п. opticus суживается, вследствие чего при открытых обоих глазах неравномерность зрачков почти совершенно исчезает. Но достаточно животному закрыть здоровый глаз и немедленно наступает расширение зрачка оперированной стороны ad maximum и вместе с тем зрачок перестает реагировать на свет.
При рассечении chiasma ηη. opticorum в передне-заднем направлении не наступает полной слепоты ни в том, ни в другом глазу. Животное, несомненно, видит и хорошо обходит препятствия, если оба глаза остаются открытыми; животное,-таким образом, почти никогда не натыкается на мебель и заворачивает еще прежде, чем достигнет стены комнаты. Глазные оси животного обыкновенно направлены прямо вдаль, зрачки большей частью не представляют резких изменений в ширине и заметно реагируют на свет, иногда не только при непосредственном раздражении светом исследуемого глаза, но и при раздражении другого глаза.
При перерезке одного из tractus opticus точно так же у животного не наблюдается полной слепоты ни в том, ни в другом глазу, и при открытых обоих глазах оно хорошо обходит препятствия. Без внимательного исследования подобного рода животное даже трудно отличить от здорового. Но достаточно завязать ему тот или другой глаз, чтобы оно начало натыкаться преимущественно на те препятствия, которые лежат в направлении неповрежденной стороны. Таким образом, при перерезке правого tractus opticus животное натыкается на препятствия, встречаемые с левой стороны, безразлично — будет ли при этом закрыт правый или левый глаз; при перерезке левого tractus, напротив того, оно наталкивается на препятствия, лежащие с правой стороны животного. Точно так же существующее ограничение поля зрения у животного легко удается обнаружить, заставляя его подбирать с полу при закрытом одном глазе разбросанные куски пищи или производя вблизи глаза угрожающие жесты рукой то с одной, то с другой стороны. В последнем случае животное обнаруживает сохранившуюся часть поля зрения миганием или стремлением отстраниться от угрожающего жеста.
,. Таким довольно сложным путем удается, наконец, убедиться, что у животного с перерезанным tractus opticus существует ограничение поля зрения в обоих глазах на стороне, противоположной в отношении к перерезанному tractus, иначе говоря — наблюдается выпадение функции одноименных отделов обеих сетчаток на сторонах, соответствующих перерезанному tractus. Мне казалось при этом, что выпадение функции в каждом глазу строго ограничивается целой половиной сетчатки, но утверждать этого с положительностью я не могу ввиду того, что более точное исследование зрительной способности у животных, вообще говоря, в высшей степени затруднительно.
Что касается ширины зрачков, то в этом отношении у животных с перерезанным tractus opticus обыкновенно не замечается никаких перемен и оба зрачка реагируют на свет вполне нормальным образом х.
Вышеприведенные результаты, таким образом, говорят с положительностью в пользу неполного перекреста волокон зрительного нерва в chiasma у собаки. Из наблюдений над животными с перерезанным tractus opticus, как мы видели, можно заключить также, что ограничение поля зрения происходит в обоих глазах и на противоположной стороне; следовательно, относительно собаки данные экспериментальных исследований вполне согласуются с гистологическими исследованиями Гуддена.
Здесь я желал бы, кроме того, обратить внимание на замеченное мною отношение зрачков при перерезке п. opticus и tractus opticus. В первом случае, как мы видели, зрачок оперированного глаза расширен и не реагирует на свет, если последний направляют непосредственно на исследуемый глаз, но он еще сокращается под влиянием света, проникающего через здоровый глаз. Между тем при перерезке одного tractus opticus, если операция произведена чисто, без посторонних повреждений, обыкновенно не замечается никакой разницы в ширине обоих зрачков, и реакция на свет в том и другом глазу остается вполне сохраненною. Эти данные, таким образом, приводят меня к заключению, что центростремительные волокна, служащие к рефлекторному сокращению зрачка, не проходят в tractus opticus, а, по всей вероятности, из п. opticus, no достижении chiasma, они проникают в самое мозговое вещество, направляясь к соответствующим ядрам п. oculomotorii.
1 Только иногда вслед за операцией в течение некоторого времени наблюдается расширение зрачка на стороне, соответствующей перерезанному нерву, которое, впрочем, скоро исчезает совершенно и. по всей вероятности, не зависит непосредственно от повреждения tractus opticus.
ОБ ОДНОЙ.'ДО СИХ ПОР НЕИЗВЕСТНОЙ СВЯЗИ МЕЖДУ ГЛАВНОЙ ОЛИВОЙ И ГОЛОВНЫМ МОЗГОМ
При исследовании мозга новорожденного примерно месячного возраста удается доказать наличие объемистого, четко отграниченного пучка миэлиновых волокон, проходящего по всему длиннику покрышки моста и мозговых ножек. Этот пучок известен, вернее, описан правильно лишь на части своего протяжения1, но его значение до настоящего времени еще неизвестно. Принимая во внимание расположение этого пучка в верхней части моста и нижней части мозговых ножек, я его хочу назвать «центральным пучком покрышки». Центральный пучок покрышки обнаруживается в продолговатом мозгу на дорзальной, гезр. наружной, стороне главной оливы, его пучки видны между ее завитками. Начиная от середины оливы, волокна центрального пучка покрышки собираются главным образом на ее дорзальной поверхности в компактный пучок, который быстро увеличивается в поперечнике; на уровне верхнего края оливы, достигнув той толщины, которая обнаруживается и выше — в мосту и мозговых ножках, он переходит в мост. Центральный пучок покрышки первоначально расположен здесь между верхней оливой и медиальной петлей (т. е. в петлевом слое минус нижняя петля); вентраль-но он отграничен трапециевидным телом. Далее кверху центральный пучок покрышки располагается более дорзально, в верхней части моста он расположен в середине сетевидной формации; еще далее кверху он все более и более приближается к центральному серому веществу полостей, к которому, наконец, прилегает на уровне верхних холмов четверохолмия, непосредственно снаружи от заднего продольного пучка. Центральный пучок покрышки на своем пути с вентральной на дорзальную поверхность покрышки переплетается с волокнами верхней мозжечковой ножки (brachium conjunctivum) и, наконец, подходит к дорзальной поверхности красного ядра, способствуя образованию дорзальной капсулы последнего. На нижних отрезках моста с центральным пучком покрышки граничит дорзально другой, меньший
] На препаратах головного мозга взрослого этот пучок также довольно легко обнаружить. У Штиллинга он очень хорошо изображен (на табл. 11, е), но ошибочно обозначен как продолжение бокового канатика. Вернике ошибочно описывает один из участков этого пучка как «пучок покрышки нз задней комиссуры».
2ft
пучок, который у плода миэлинизируется гораздо раньше, у новорожденного же, примерно месячного и более старшего возраста, он представляется частично слившимся с центральным пучком покрышки. Этот пучок очень резко выделяется от остальных отделов сетевидпой формации, особенно у семимесячного плода, и, таким образом, выходит, что он не имеет никакой связи с центральным пучком покрышки. Напротив того, он состоит из волокон, которые исходят из серых масс боковых отделов ромбовидной ямки на уровне вхождения тройничного нерва, дугообразно вступают в сетевидную формацию, далее идут косо книзу к одноименной главной оливе и пронизывают ее сзади и снаружи вперед и внутрь, исчезая в межоливном слое петли (formatio reticularis).
Там, где пучки вентральной части г задней комиссуры мозга идут к заднему продольному пучку, они частично переплетаются с центральным пучком покрышки, так что оба пучка кажутся как бы переходящими друг в друга (отсюда и ошибка Вернике). Однако изучение плода примерно семимесячного возраста, у которого в это время указанные волокна задней комиссуры обладают толстыми миэлиновыми оболочками, в то время когда центральный пучок покрышки еще совершенно лишен миэлина, показывает, что оба пучка не связаны друг с другом. Больше того, центральный пучок покрышки продолжается дальше кверху по направлению к большому мозгу, в котором он между дном III желудочка и красным ядром, в некотором отдалении от первого, загибается вперед, resp. вентрально; я, однако, не могу указать место его центрального окончания.
В пользу того, что центральный пучок покрышки в периферических отделах связан с главными оливами, говорит следующее: центральный пучок покрышки получает миэлиновые оболочки непосредственно после той системы волокон, которая соединяет перекрестно большие оливы с мозжечком (главная часть верев-чатого тела), т. е. в последний месяц утробной жизни. Только к этому времени миэлинизируются и пучки волокон, исчезающие в сером веществе олив. Также можно с достоверностью исключить переход центрального пучка покрышки в спинной мозг. Кроме того, поперечное сечение центрального пучка покрышки и пучка, осуществляющего соединение мозжечка с большой оливой, примерно одинаково.
Центральный пучок покрышки особенно хорошо выделяется в мосту как раз у новорожденного месячного возраста. Это связано с тем, что проходящие медиально и латерально от центрального пучка покрышки довольно объемистые пучки волокон покрышки или совершенно лишены миэлина, или бедны^им; лишь позже они становятся богатыми миэлином.
1 Ср. Darkschewitsch, Ueber die hintere Commissura des-Gehirns, Neurol. Zbh, 1885, λη<>5.
О СОЕДИНЕНИЯХ ВЕРХНИХ ОЛИВ И ОБ ИХ ВЕРОЯТНОЙ ФИЗИОЛОГИЧЕСКОЙ РОЛИ
Верхние оливы представляют собой образование, соединения которых с другими частями центральной нервной системы до последнего времени оставались почти совершенно неизвестными или по крайней мере невыясненными в достаточной степени. Это обстоятельство, без всякого сомнения, зависело в значительной степени от того, что верхние оливы расположены в се-тевидной формации, среди волокон которой нелегко проследить действительные их соединения.
При исследовании мозгов зародышей я убедился, что волокна, выходящие из верхних олив, представляются мякотными еще в очень раннем возрасте, а именно у зародышей 28—30 см длиной, приблизительно на 5—6-м месяце утробной жизни, когда большинство волокон стволовой части мозга, а также и волокна наружного отдела сетевидной формации еще совершенно безмя-котны.
Ввиду этого мозги зародышей указанного возраста являются лучшим материалом для изучения хода волокон, соединяющих верхние оливы с другими образованиями стволовой части мозга.
Исследование таких мозгов показывает, что соединения верхних олив весьма обширны и разнообразны. Прежде всего в них берут начало два больших пучка, из которых один поднимается кверху, к области заднего четверохолмия, а другой направляется в мозжечок, входя в состав внутреннего отдела задней его ножки.
Первый пучок представляет собой не что иное, как пучок, известный под названием нижней петли, который, однако, по моему мнению, было бы правильнее назвать боковой петлей *. На основании исследования мозгов, принадлежащих плодам в 28—30 см длиной, у которых из всех частей петельного слоя
1 См. мое сообщение «Untersuchungen iiber die Schleifenschicht» в Sit-zungsbericht math.- phys. Klasse der Konigl. Sachs. Gesellschaft der Wis-senschaften, 1885.
^Schleifenschicht немцев) мякотными являются только волокна боковой или нижней петли, можно утверждать с положительностью, что эти последние волокна не происходят из межолив-чатого слоя, как волокна других частей петельного слоя,.а обракуют собой совершенно самостоятельный пучок, обнаруживающийся впервые только на уровне трапециевидного тела.
Волокна этого пучка, как я убедился, берут начало главным образом в сером веществе верхней оливы соответствующей стороны; некоторые же из них, входя в состав поперечных волокон трапециевидного тела, повидимому, происходят и из противоположной верхней оливы.
В направлении кверху от трапециевидного тела волокна боковой петли, отодвигаясь постепенно кнаружи вместе с верхним концом верхней оливы, вскоре поворачивают к заднему четверохолмию; затем большинство их оканчивается в ядре последнего, а меньшая часть рассеивается в окружающем сером веществе. Несомненно, что волокна боковой петли не переходят в область переднего четверохолмия, как это допускалось некоторыми авторами1.
Второй пучок, поднимающийся от верхних олив к мозжечку, был уже описан мною в статье «О двух пучках, входящих в состав внутреннего отдела задней ножки мозжечка»2. Он располагается между волокнами наружного отдела мозжечковой ножки и особенным пучком, выходящим к мозжечку из области описанного мною скопления клеточных элементов, расположенного непосредственно позади наружного слухового ядра, или ядра Дейтерса. По вступлении в мозжечок волокна рассматриваемого пучка, берущего начало главным образом в соответствующей, а частью, быть может, и в противоположной верхней оливе, проходят над передней ножкой мозжечка и направляются затем к кровельным ядрам как соответствующей, так и противоположной стороны.
Кроме двух только что описанных пучков, принадлежащих верхним оливам, на разрезах из мозгов зародышей удается еще рассмотреть отношение этих образований, во-первых, к переднему ядру слухового нерва (nucleus anterior Meynerti, nucleus acustici «accessorius» Schwalbi), во-вторых, к ядру отводящего
1 Необходимо заметить, что волокна нижней петли на весьма значительном протяжении кверху сопровождаются группой клеточных элементов, исчезающих только в области заднего четверохолмия, на небольшом расстоянии от его ядра. Так как при этом на непрерывном ряде разрезов не удается найти свободного промежутка между этой группой клеточных элементов и соответствующей верхней оливой, то первую и следует, по моему мнению, рассматривать как прямое продолжение последней. Верхние оливы, таким образом, заходят кверху значительно далее, чем считали до настоящего времени.
2 Врач, 1885, стр. 131; Neurol. Zbl., 1885, № 7.
32
нерва1 и, в-третьих, к волокнам основного пучка боковых столбов спинного мозга2.
волокна, связывающие верхние оливы с передним ядром слухового нерва, выходят из последнего в поперечном направлении наперекрест с передним корешком слухового нерва и рассеиваются затем в соответствующей верхней оливе. Часть этих волокон, впрочем, переходит и в поперечные волокна трапециевидного тела, направляясь к противоположной оливе.
Соединение верхних олив с ядром отводящего нерва на препаратах из мозгов зародышей, окрашенных гематоксилином по Вейгерту, видно с поразительной наглядностью. Волокна, при посредстве которых происходит это соединение, поднимаются от оливы к ядру отводящего нерва в виде довольно широкого пучка, проходящего почти параллельно нисходящей части корешка лицевого нерва. Повидимому, однако, не все эти волокна оканчиваются в самом ядре отводящего нерва: некоторая часть их переходит и в область заднего продольного пучка, направляясь, быть может, к ядрам глазодвигательного нерва.
Наконец, что касается до отношения верхних олив к основному пучку боковых столбов спинного мозга, то его без труда удается показать на препаратах из мозга зародышей приблизительно в 25—28 см длиной, у которых из всех частей белых столбов спинного мозга представляются мякотными только корешковая область пучков Бурдаха и основной пучок передних и боковых столбов. Делая разрезы из таких мозгов, мы убеждаемся, что при переходе спинного мозга в продолговатый самая задняя часть основного пучка боковых столбов отделяется от других волокон этого пучка, переходящих во внутренние части сетевид-ной формации, и затем в виде обособленного пучка поднимается по периферии продолговатого мозга, проходя между нижними оливами и восходящим корнем тройничного нерва. На непрерывном ряде разрезов легко доказать, что этот пучок продолжается кверху только до области верхних олив, в которых он и оканчивается3.
Вышеописанные соединения верхних олив позволяют нам сделать некоторые заключения об их отправлении, оставшемся до настоящего времени еще совершенно неизвестным.
1 На отношение волокон трапециевидного тела (происходящих главным образом из верхних олив) к переднему ядру слухового нерва указал еще Флексиг в своей работе «Die Leitungsbahnen im Gehirn und Rflcken-mark»(Leipzig,1876); соединение же верхних олив с ядром отводящего нерва описывалось уже Дином, но все другие авторы о нем не упоминают совсем.
2 Основным пучком бокового столба я называю ту часть остаточной области последнего (Seitenstrangrest Флексига), которая остается за вычетом из нее пограничной области (Grenzschicht der grauen Substanz)n описанного мною периферического пучка передней области бокового столба, облагающегося миэлином в относительно позднем возрасте (см. протоколы заседаний Общества психиатров в С.-Петербурге за ноябрь 1884 г.).
3 Непосредственное продолжение его в боковую петлю невероятно уже и ввиду того обстоятельства, что волокна последней облагаются миэлнном несколько позже, чем волокна рассматриваемого пучка.
^ В. М. Бехтерев
33
Обширная связь этих образований с другими гнездами серого вещества стволовой части мозга говорит, очевидно, в пользу того, что верхние оливы играют роль важного отражающего центра. В этом отношении в особенности обращает на себя внимание соединение верхних олив с ядром отводящего нерва—соединение, указывающее на отношение этих образований к отраженному передвижению глазных яблок. Ввиду этого получает большое значение и связь верхних олив с наружным ядром слухового нерва. Соединение это, очевидно, может быть рассматриваемо как проводящий путь, благодаря которому слуховые впечатления могут вызывать отраженные движения глазных яблок. Подобное же значение в отношении отраженной передачи зрительных импульсов к верхним оливам, повидимому, следует приписать и соединение последних с задним четверохолмием при посредстве волокон боковой петли.
Что касается соединения верхних олив с мозжечком, то оно может объяснить нам происхождение тех отраженных расстройств в положении и передвижении глазных яблок, которые обычно наблюдаются при электризации и разрушениях мозжечка.
Наконец, упомянутое выше отношение верхних олив к задней части основного пучка боковых столбов (волокна которого, как известно, стоят в непосредственной связи с клеточками передних рогов) дает право заключить, что физиологическая роль этих образований не ограничивается отраженным передвижением глазных яблок, а имеет некоторое отношение и к другим мышечным группам нашего тела.
Какие движения при этом имеются в виду, сказать с точностью, разумеется, нельзя. Можно полагать, однако, что одновременно с движением глазных яблок верхние оливы управляют и отраженным движением головы. В пользу этого предположения говорит по крайней мере то обстоятельство, что упомянутые движения представляются тесно сочетанными между собой и составляют одновременный результат влияния слуховых и зрительных впечатлений.
О ДВУХ ПУЧКАХ, ВХОДЯЩИХ В СОСТАВ ВНУТРЕННЕГО
ОТДЕЛА ЗАДНЕЙ НОЖКИ МОЗЖЕЧКА И О РАЗВИТИИ
ВОЛОКОН СЛУХОВОГО НЕРВА
При исследовании мозгов зародышей я убедился, что во внутреннем отделе задней мозжечковой ножки поднимаются к мозжечку по крайней мере два отдельных пучка. Один из этих пучков облагается миэлином в очень раннем возрасте — у зародышей приблизительно в 28—30 см длиной; другой же, располагающийся кнутри от первого, становится мякотным только у зародышей приблизительно в 35—38 см.
Последний пучок берет свое начало в особом скоплении небольших клеточных элементов, расположенном позади наружного слухового ядра, или ядра Дейтерса (частью же, может быть, и в самом ядре Дейтерса). Поднимаясь во внутренней части мозжечковой ножки, в соседстве с наружной стенкой IV желудочка, рассматриваемый пучок, вступив в мозжечок, частью проходит между волокнами передней ножки мозжечка, а частью идет над ней и рассеивается затем между серыми ядрами шарообразного и пробковидного тела (Kugelkern и Pfropf немцев). Несомненно при этом, что волокна его не переходят на другую сторону, и, следовательно, не служат к образованию перекреста по срединной линии над кровельными ядрами.
Другой пучок внутреннего отдела мозжечковой ножки проходит тотчас снаружи и частью впереди от первого. Таким образом, он поднимается между вышеописанным пучком и волокнами прямого мозжечкового пучка, выходящего из боковых столбов спинного мозга, огибает затем снаружи и сверху переднюю ножку мозжечка и направляется к кровельным ядрам. Значительная часть этого пучка образует над упомянутыми ядрами, частью же в промежутке между ними по срединной линии описанный Мей-нертом перекрест с волокнами подобного же пучка, идущего с противоположной стороны, и затем оканчивается в противоположном кровельном ядре; небольшая часть волокон этого пучка оканчивается, повидимому, в соответствующем кровельном ядре.
Что касается происхождения этого пучка в продолговатом мозгу, то я убедился, что часть его волокон берет начало в верхних оливах"
3*
Некоторые из волокон этого пучка, повидимому, имеют отношение только к противоположной оливе, так как, вступая в состав поперечных волокон трапециевидного тела, они огибают соответствующую оливу и переходят затем в противоположную сторону. Но во всяком случае значительная его часть, несомненно, берет начало в соответствующей верхней оливе. По крайней мере на многих препаратах удается видеть довольно значительное число волокон, выходящих из соответствующей верхней оливы, которые, проникая частью через студневидное вещество (substantia gelatinosa) между волокнами восходящего корня тройничного нерва, частью же проходя снаружи от последнего, продолжаются затем в пучке, поднимающемся к кровельным ядрам мозжечка.
Необходимо заметить еще, что некоторые из волокон упомянутого пучка не переходят в трапециевидное тело, а, поворачивая кнутри тотчас при основании ядра Дейтерса, вскоре исчезают на пространстве между последним и восходящим корнем тройничного нерва.
Слуховой нерв по развитию своих волокон может быть разделен на два отдельных корня: один, соответствующий переднему корешку авторов, представляется мякотным уже у зародышей в 25 см длиной, тогда как другой, соответствующий заднему корешку авторов, облагается впервые миэлином только у зародышей приблизительно в 30 см. Первый из этих корешков происходит из предцверной части слухового нерва и потому может быть назван корешком преддверного нерва; второй же происходит из улитковой ветви слухового нерва, вследствие чего может быть назван корешком улиткового нерва.
Ни один из этих корешков не имеет прямого соединения с мозжечком. Корешок п. vestibularis большинством своих волокон оканчивается в упомянутом выше скоплении клеточек, расположенном непосредственно сзади ядра Дейтерса; другая же часть его волокон спускается вдоль последнего к продолговатому мозгу, сопровождаясь на некотором протяжении волокнами ранее развивающегося пучка внутреннего отдела задней мозжечковой ножки. Корешок же п. cochlearis оканчивается значительной частью своих волокон в так называемом переднем ядре слухового нерва (nucleus anterior, по Мейнерту, nucleus acustici acces-sorius, no Швальбе).
О других соединениях я сообщу позднее. Здесь же следует упомянуть еще, что волокна striae acusticae облагаются миэлином много позднее обоих корешков слухового нерва, вследствие чего они и не могут представлять собою прямого продолжения последних.
О ЗАДНИХ КОРЕШКАХ, МЕСТЕ ИХ ОКОНЧАНИЯ
В СЕРОМ ВЕЩЕСТВЕ СПИННОГО МОЗГА
И ОБ ИХ ЦЕНТРАЛЬНОМ ПРОДОЛЖЕНИИ
Имея значительное количество препаратов мозгов человеческих зародышей и новорожденных, я поставил себе, между про^ чим, целью выяснить на этом материале окончание задних корешков и их дальнейшее распределение внутри серого и белого вещества спинного мозга. В этом направлении исследования мои начались еще в течение зимы 1884/85 г.; предварительное же сообщение о результатах этих исследований было сделано мною в начале 1885 г. в заседании Психиатрического общества1. Кроме того, позднее была напечатана мною относящаяся сюда же работа: «Об особенной составной части боковых столбов спинного мозга»'2. До сих пор, однако, мне не удалось опубликовать свои исследования по этому предмету в более полном объеме, чем я и желал бы оправдать появление настоящей работы.
За указанный промежуток времени были обнародованы три исследования, относящиеся к данному предмету: подробная работа Лиссауэра3 и краткие сообщения Такакса4 и Россолимо5. Эти работы я считаю здесь необходимым прореферировать, прежде чем перейти к изложению данных, полученных при своих исследованиях.
Лиссауэр, занимавшийся исследованием патологоанатомиче-ских изменений при tabes dorsalis, приводит в своей работе подробное описание анатомического отношения задних корешков
1 См. протоколы этого общества за 1885 г.
* См. Врач за 1885 г. и Du Bois-Reymond's Arch., Anat. Abt., 1886. 3 Lissauer, Beitrag zum Faserverlauf im Hinterhorn d. mensch-
lichen Rfickenmarks etc.. Arch. f. Psych, u. Nervenkrankh., Bd. XVII, Hft. 2 (ранее этой работы Лиссауэ'ром были опубликованы краткие сообщения в Neurol. Zbl., 1885, № 11, и в Verhandlungen des IV Kongresses f, innere Medizin zu Wiesbaden).
* T a k а с s, Ueber d. Verlauf d. hinteren Wurzelfasern im Ruckenmark etc., Neurol. Zbl., 1887, № 1.
8 Россолимо, Zur Frage uber den weiteren Verlauf der Hinter-wurzelfasern im Rflckenmarke, Ibid., 1886, № 17. После того как настоящая статья была приготовлена к печати, появилась уже и подробная работа Россолимо в виде диссертации: «Экспериментальное исследование по вопросу о путях, проводящих чувствительность и движение», М., 1887.
37
в поясничном утолщении спинного мозга, следовательно, в той части последнего, в которой преимущественно и локализуется патологический процесс в начальном периоде tabes.
При этих исследованиях автор убедился, что тонкие, вертикально поднимающиеся волокна в окружности наружного края желатинозного вещества заднего рога или в краевом поясе (resp. в описанной мною заднекорешковой части боковых столбов1) принадлежат задним корешкам. Эти волокна, по исследованиям Лиссауэра, большей частью переходят в желатинозное вещество заднего рога.
Не касаясь подробно рассматриваемого автором вопроса о строении желатинозного и спонгиозного вещества заднего рога, мы остановимся здесь только на результатах его относительно содержания в последнем задних корешков.
Задний рог, по автору, содержит корешковые волокна двух родов: 1) толстые волокна, проникающие пучками непосредственно до substantia spongiosa и здесь изменяющие свое горизонтальное направление на вертикальное; 2) тонкие волокна, которые тотчас по вступлении в спинной мозг отклоняются кнаружи, образуя собой восходящие волокна краевого пояса (Randzone).
Здесь эти волокна проходят на большем или меньшем протяжении вдоль спинного мозга и затем вступают в задний рог. Дальнейшее же распределение этих волокон в сером веществе спинного мозга остается еще во многих отношениях невыясненным.
Что касается исследований Такакса, то они опубликованы только в виде коротких положений, часть из которых я приведу здесь дословно.
1. Задние корешки при вступлении в спинной мозг разделяются на две части. Одна часть проникает в серое вещество, тогда как другая загибает внутрь и кнаружи в белое вещество, окаймляющее задние серые рога.
2. Корешковые волокна, вступающие в серые рога, проникая через substantia gelatinosa, достигают клеток кларковых столбов и, повидимому, в них предварительно оканчиваются.
3. Корешковые волокна, вступающие в белое вещество, окаймляющее серые задние рога (клиновидный пучок Бурдаха и задний отдел остаточной области боковых столбов), загибаются тотчас кверху, поднимаются на нем по крайней мере на высоту трех корешков и затем входят в вещество заднего рога.
Из кларковых столбов, по автору, часть волокон направляется во внутренние области пучков Бурдаха, образуя здесь крыловидную часть разреза (fliigelformigen Flachenabschnitt), из которой позднее создается пучок Голля; другая же часть волокон из кларковых столбов, направляющаяся к периферии боковых столбов, образует собой мозжечковый пучок.
1 В последующем изложении мы будем называть эту область «наружной заднекорешковой областью» в отличие от «внутренней заднекорешковой области», или корешковой области пучков Бурдаха.
38
Как пучок Голля, так и мозжечковый пучок боковых столбов составлены, по автору, из однородных волокон, служащих при посредстве клеток заднего рога продолжением задних корешков. Пучки же Бурдаха и задний отдел остаточной области боковых столбов образованы преимущественно из непосредственного продолжения задних корешков, которые на большем или меньшем расстоянии выше места вхождения вступают в серое вещество заднего рога. Автор допускает также, что в упомянутых частях спинного мозга к корешковым волокнам присоединяются еще волокна, связывающие между собой различные уровни серых задних столбов.
Следует еще заметить здесь, что оба цитированные автора делают из своих исследований соответствующие выводы относительно патологии tabes dorsalis, подкрепляя их патологоана-томическими исследованиями.
Относительно работы Россолимо необходимо упомянуть, что на основании опытов с перерезками задних корешков у морской свинки он доказывает, что пучки Голля не служат непрерывным продолжением волокон задних корешков — положение, которое было высказано мною еще раньше на основании исследований с помощью метода развития (см. цитированное выше сообщение)1.
Мои исследования, как было уже упомянуто, произведены на мозгах зародышей и младенцев различного возраста, причем препараты окрашивались преимущественно гематоксилином по Вейгерту, частью же хлористым золотом по Фрейду. Результаты, полученные при этих исследованиях, я постараюсь изложить здесь в возможно сжатой форме.
При исследовании мозгов зародышей прежде всего выяснилось, что задние корешки составлены не из однородных волокон. Одни из них развиваются очень рано, представляясь мякотными уже к началу пятого месяца утробной жизни плода (у зародышей 25 см длиной), тогда как другие облагаются миэлином много позднее, незадолго до времени рождения.
Рис. 1.
1 В более подробной работе о том же предмете (loc. cit.) автор указывает на следующие изменения вслед за упомянутой операцией: перерождение корешковых волокон, входящих в задний рог, атрофию большинства клеток этого рога, уменьшение объема заднего столба соответствующей стороны за счет наружных его волокон, атрофию пояса, прилежащего к периферии заднего рога, уменьшение количества волокон в заднем углу соответствующего бокового столба и разрежение сети в переднем роге той же стороны.
39
Таким образом, задние корешки по времени развития волокон могут быть разделены по крайней мере на два строго диференци-рованных пучка. Волокна этих пучков на поперечном разрезе заднего корешка не выделены отдельно, а смешаны между собой более или менее равномерно. В спинном же мозгу оба упомянутые пучка имеют различное анатомическое распределение. Так, большинство волокон ранее развивающегося пучка, как это видно из рис. 1 (rpi), тотчас по вступлении в спинной мозг отклоняется кнутри и вступает в корешковую область пучков Бурдаха, меньшая же часть проникает непосредственно в substantia gelatinosa, преимущественно в ее внутренние, а отчасти (преимущественно в нижних областях спинного мозга) и в наружные области. Между тем большинство волокон позднее развивающегося пучка (гре) при вступлении в спинной мозг поворачивает кнаружи, входя в наиболее заднюю часть боковых столбов, и здесь поднимается на некотором протяжении кверху; меньшая же часть или входит непосредственно в substantia gelatinosa, или поворачивает кверху уже непосредственно по вступлении в спинной мозг, располагаясь между корешковыми волокнами ранее развивающегося пучка. В нижних областях спинного мозга (в поясничном утолщении и в нижнем отделе грудной области) небольшая часть рассматриваемого пучка отклоняется даже кнутри от последних, занимая самую наружную часть пучков Бурдаха.
Тот и другой пучок различаются между собой не только по времени развития, но и по калибру волокон: первый, т. е. ранее развивающийся пучок, состоит из волокон значительной толщины, тогда как позднее развивающийся пучок содержит сравнительно очень тонкие волокна. Для краткости в последующем изложении первый пучок мы будем называть внутренними толстыми корешками, а второй — наружными тонкими корешками.
Следует, впрочем, иметь в виду, что название «внутренние и наружные корешки» в данном случае не вполне соответствует действительности, так как волокна первых, как упомянуто, заходят частью и в наружные области substantiae gelatinosae, а волокна вторых частью располагаются даже кнутри от первых, занимая самую наружную, прилежащую к корешкам область пучков Бурдаха1.
Вопреки распространенному мнению, мы считаем, что все волокна задних корешков, вступая рано или поздно в серое вещество спинного мозга, прерываются содержащимися здесь клеточ-
1 Это замечание имеет силу главным образом по отношению к поясничной части спинного мозга; в других же областях последнего топография того и другого пучка задних корешков более или менее полно соответствует их названию. Нелишне еще заметить, что мое разделение на внутренние толстые и наружные тонкие корешки, как легко видеть из описания, не вполне соответствует общепринятому в учебниках анатомии делению зад-, них корешков на внутренний наружный пучок.
40
ными элементами. Противоположное мнение, как известно, высказывалось нередко относительно внутреннего пучка задних корешков, часть волокон которого будто бы непрерывно поднимается в задних столбах, именно в пучке Голля, к продолговатому мозгу.
Это мнение основывалось главным образом на том обстоятельстве, что при перерезке п. ischiadic! у животных и при сдавлении caudae equinae у человека с течением времени обнаруживается вторичное перерождение задних столбов, причем перерождение пучков Голля распространяется вверх до продолговатого мозга1.
Подобные случаи, однако же, по моему мнению, ничуть не могут служить доказательством непрерывного продолжения задних корешков к продолговатому мозгу. Последнее очевидно, по крайней мере, из того обстоятельства, что вторичное перерождение волокон далеко не всегда оканчивается в сером веществе, в элементах которого прерываются перерождающиеся волокна; напротив того, весьма нередко случается, что за перерождением определенной системы волокон следует перерождение или атрофия элементов серого вещества, а затем и перерождение волокон, лежащих по другую сторону клеточных элементов. Известно, например, что перерождение передней ножки мозжечка обыкновенно влечет за собой атрофию красного ядра и принадлежащих ему волокон; при повреждении мозговой коры (лобных извилин) наблюдали перерождение венечных волокон зрительного бугра, атрофию его переднего ядра и перерождение выходящего из последнего к corpus mamillare пучка Вик-д'Азира; наконец, перерождение пирамидных пучков боковых столбов спинного мозга в некоторых случаях, как известно, вызывает последовательную атрофию клеток передних рогов и перерождение волокон передних корешков.
В пользу того, что задние столбы спинного мозга не могут быть рассматриваемы как непрерывное продолжение волокон задних корешков к продолговатому мозгу, говорит с очевидностью тот факт, что волокна пучков Голля и большинство волокон периферической части пучков Бурдаха облагаются миэлином много позднее волокон внутренних толстых корешков. Только передне-наружная часть пучков Бурдаха становится мякотной одновременно с волокнами внутренней части задних корешков и, очевидно, составляется по преимуществу из корешковых волокон2. Последние, однако, в этой части задних столбов, как показывают точные гистологические исследования, поднимаются кверху только на незначительном протяжении и затем, снова
1 Что это перерождение ничуть не может считаться обязательным, по крайней мере для некоторых животных (морских свинок), доказывается недавними исследованиями Россолимо (см. выше).
2 О разделении пучков Бурдаха на передне-наружную, или корешковую, и заднюю, или периферическую, части см. мою работу «О составе задних столбов спиниого мозга», Врач, 1885, и Neurol. Zbl., 1885, № 2.
41
принимая горизонтальное направление, рано или поздно вступают в серое вещество спинного мозга.
Что касается места окончания волокон задних корешков, то изучение препаратов из мозгов новорожденных и зародышей убеждает нас в следующем.
Большинство волокон внутренних толстых корешков, как упомянуто, вступает в передне-наружную часть пучков Бурдаха или внутреннюю корешковую область (rpi'), меньшая же часть проникает непосредственно в substantia gelatinosa Rolandi. В передне-наружной части пучков Бурдаха многие из корешковых волокон загибаются кверху, после чего они снова принимают горизонтальное направление, устремляясь к серому веществу заднего рога. Другая же часть корешковых волокон, обогнув выдающуюся кнутри часть заднего рога, проникает прямо в серое вещество последнего.
По вступлении в серое вещество заднего рога волокна внутренних толстых корешков получают двоякое направление.
Одни из них, располагающиеся более кнутри и проходящие через передне-наружную область пучков Бурдаха в виде компактного пучка, подходят к серым кларковым столбам (kl) и здесь рассеиваются между клеточными элементами последних; в областях же спинного мозга, где отсутствуют кларковы столбы, этот пучок оканчивается в соответствующих ядрах шейной и сакральной части спинного мозга. Напротив того, более наружные волокна внутренних толстых корешков из передне-наружной части пучков Бурдаха, прямо или временно изменяя свое направление на вертикальное, направляются впереди роландова вещества (загибаясь кверху или книзу) далее вглубь серого вещества. Некоторые из волокон этого пучка теряются уже в средней области серого вещества, вступая в соединение с расположенными здесь клеточными элементами, тогда как другие волокна достигают его передних рогов и здесь входят в состав сети нервных волокон, раскинутой между тремя большими группами клеток. Наконец, часть волокон этого пучка задних корешков по достижении средней области серого вещества поворачивает к передней комиссуре и, присоединяясь к ее волокнам, переходит в основной пучок противоположного переднего столба, а оттуда в передний рог другой стороны (см. рис. 1).
Что касается позднее развивающихся наружных тонких корешков, то исследование мозгов новорожденных младенцев показывает, что эти корешки с самого начала поднимаются на некотором протяжении кверху внутри задней части боковых столбов (в пучке, названном мною наружной задне-корешковой областью) или в краевом поясе Лиссауэра {гре') и затем снова вступают в серое вещество заднего рога. Проходя через наружные отделы sub-stantiae gelatinosae, частью же по наружному краю ее, эти волокна прерываются в небольших клетках заднего рога, располагающихся частью рассеянно, частью более или менее скученно
42
тотчас впереди роландова вещества (см. рис. 1). Небольшая часть этих волокон, повидимому, направляется также вглубь •серого вещества, подходя к боковой группе клеток переднего рога.
Таким образом, волокна того и другого пучка задних корешков имеют крайне разнообразное распределение в сером веществе спинного мозга; они вступают в соединение почти со всеми более или менее значительными группами клеток последнего и, кроме того, при посредстве передней комиссуры переходят на другую сторону спинного мозга.
Из кларковых столбов, служащих местом окончания значительной части толстых внутренних корешков, в свою очередь возникают многочисленные волокна, устанавливающие соединение их серого вещества с другими областями центральной нервной системы. От кларковых столбов эти волокна расходятся по трем главным направлениям. Прежде всего в них берут начало волокна, служащие к образованию мозжечкового пучка боковых столбов (Флексиг). Эти волокна выходят с передней и частью наружной стороны кларковых столбов и, заворачивая затем дугообразно к наружным отделам серого вещества (между задним и боковым рогом), переходят через белые боковые столбы, направляясь к периферии задней их части. В особенно обильном количестве эти волокна встречаются в области перехода поясничного утолщения в грудную часть спинного мозга, где их и удается лучше всего исследовать. Несомненно, впрочем, что волокна, •образующие прямые мозжечковые пучки (fc), выходят из кларковых столбов и на более высоких уровнях спинного мозга, хотя и «в значительно меньшем количестве.
Другие волокна выходят также в виде довольно значительных •пучков с задней и частью внутренней стороны кларковых столбов во внутренние и задние части пучков Bypnaxa(fpe). Небольшая часть этих волокон, по всей вероятности, переходит и в пучки Голля (fpi), в которых среди поздно развивающихся волокон мы всегда находим известное количество волокон, облагающихся ми-элином одновременно с волокнами периферической или задней части пучков Бурдаха. Несомненно, однако, что большая часть волокон пучков Голля, развивающаяся много позднее волокон пучков Бурдаха, берет начало не из кларковых столбов, а, как увидим впоследствии, из рассеянных клеток задних рогов.
Наконец, третьи волокна выходят с передней стороны кларковых столбов и, направляясь с самого начала почти прямо вперед, частью продолжаются затем в область соответствующего переднего рога, частью заворачивают в область передней комиссуры (са), вместе с волокнами которой они переходят на другую сторону спинного мозга (в противоположный передний рог).
Те и другие из только что упомянутых волокон удается встретить почти на всех з'ровнях грудной части спинного мозга, хотя
43
в более низких ее отделах они встречаются, повидимому, в большем количестве, нежели в верхних.
Из мелких клеток задних рогов, частью рассеянных, частью скученных непосредственно впереди роландова вещества и служащих местом окончания наружных тонких корешков, возникают волокна также тонкого калибра, которые, огибая кларковы столбы изнутри, а частью и снаружи, проходят в серую спайку (как в ее задний, так и в передний отдел — commissura anterior et posterior grisea). Из последней, как показывает сравнение большого ряда разрезов, они направляются в боковые столбы спинного мозга в область пограничного слоя, где образуют особую систему волокон, лежащих более или менее рассеянно между волокнами задней части основного (ff) и внутренней части пирамидного пучков (fpl).
Из рассеянных клеток заднего рога берет свое начало также и большинство волокон пучков Гол ля (jpi), в чем легко убедиться на разрезах из сакральной области спинного мозга.
Волокна, служащие для образования последних, проходят, подобно предыдущим, по серому веществу заднего рога в ближайшем соседстве с внутренним краем последнего; внутри задней комиссуры вблизи срединной линии (но не переходя через нее) они круто поворачивают почти прямо кзади и преходят затем вблизи задней перегородки до периферических областей срединной части задних столбов, где впервые и обозначаются пучки Голля1.
Таким образом, как внутренние толстые, так и наружные тонкие задние корешки (не говоря о волокнах, идущих непосредственно к клеткам средней части серого вещества и передних рогов и имеющих, по всей вероятности, значение рефлекторных связей) служат для образования двух самостоятельных систем: первые дают начало волокнам задне-внутренней, или периферической, части пучков Бурдаха и мозжечковым пучкам боковых столбов, вторые служат для образования волокон особой системы в противоположном боковом столбе и волокон внутреннего пучка задних столбов или пучка Голля соответствующей стороны.
Несомненно, что два пучка задних корешков, развивающиеся в различное время утробной жизни плода, должны иметь и различное функциональное значение. До сих пор, однако, мы не имеем еще достаточно данных, чтобы решить окончательно вопрос о функциональном значении того и другого пучка задних кореш-
1 Часто высказываемое мнение, что пучки Голля впервые образуются в поясничном утолщении спинного мозга, я считаю ошибочным. Исследование мозгов зародышей раннего возраста, где пучки Голля представляются еще безмякотными, в то время как другие части задних столбов уже обложились миэлином, показывает с несомненностью, что эти пучки находятся и ниже поясничного утолщения, в сакральной части спинного мозга, в виде двух узких клиньев, расположенных по обе стороны задней перегородки.
ков. Но одно обстоятельство, по моему мнению, заслуживает в указанном отношении полного внимания: при опытах с перерезкой задних столбов спинного мозга у различных животных меня всегда поражал тот факт, что повреждения, несомненно, касавшиеся корешковой области пучков Бурдаха, не вызывали у жи- „ вотных криков и, следовательно, ощущений. Последние обнаруживались только в том случае, когда повреждались части спинного мозга, лежащие при apex cornu posterioris или же когда режущий инструмент продвигался слишком далеко вперед и вместе с задними столбами повреждал серое вещество задней спайки или основания задних рогов.
На основании этих данных можно предполагать, что для проведения кожной чувствительности служат наружные тонкие, а не внутренние толстые корешки. Роль же последних, быть может, кроме рефлекторной функции, ограничивается проводимостью мышечного чувства.
В пользу этого последнего предположения, мне кажется, говорят до известной степени патологоанатомические данные при tabes dorsalis, когда процесс в самом начале ограничивается нередко поражением одной передне-наружной или корешковой области пучков Бурдаха (содержащей волокна внутренних толстых корешков), а начальным симптомом является всегда более или менее значительное поражение мышечного чувства.
Если наши предположения верны, то спрашивается, какие из двух систем, являющихся центральным продолжением той или другой части задних корешков, должны служить проводниками обоих родов чувствительности?
Из двух систем, принадлежащих внутренней части задних корешков мозжечкового пучка и периферической части пучков Бурдаха, первый не может служить проводником мышечных ощущений на том основании, что он поднимается непосредственно в мозжечок, поражение которого (и даже почти полное удаление у животных) не нарушает мышечной чувствительности. Следует, таким образом, полагать, что проводником последней является задняя, или периферическая, область пучков Бурдаха, мозжечковый уже пучок имеет, очевидно, рефлекторное значение; по нему, по всей вероятности, проходят центростремительные импульсы к мозжечку, участвующие при выполнении функции равновесия.
Что касается проведения кожной чувствительности, то из. двух систем, служащих продолжением наружного пучка задних корешков и заложенных — одна в соответствующем пучке Голля, другая в противоположном боковом столбе,— первая не может быть принята в расчет: 1) ввиду отрицательных физиологических и патологических данных и 2) ввиду того что эта система волокон не содержит перекреста в спинном мозгу, тогда как все физиологические данные говорят за существование такого перекреста для чувствующих волокон.
45
Таким образом, проводником кожных ощущений следует считать систему волокон, заложенную в боковых столбах; пучки же Голля, подобно мозжечковым, мы рассматриваем как пучки, имеющие рефлекторное значение.
Ввиду связи пучков Голля при посредстве ядер в продолговатом мозгу с мозжечком и через петельный слой с описанным-мною сетчатым ядром покрышки и зрительным бугром, следует думать, что эти пучки служат частью, подобно мозжечковым пучкам, для поддержания равновесия тела, частью же для сложных рефлексов, вызываемых раздражением кожной поверхности тела.
В заключение два слова о задней комиссуре спинного мозга.
В тех мозгах зародышей, в которых наружные тонкие корешки, а равно и пучки Голля еще совершенно безмякотны, задняя комиссура также не содержит еще ни одного миэлинового волокна. Очевидно, следовательно, что задняя комиссура вовсе не содержит волокон, входящих в состав внутренних толстых корешков, развивающихся, как мы видели, в очень раннем возрасте.
Только у зародышей незадолго перед рождением и у новорожденных мы впервые встречаем в задней комиссуре нежные мякот-ные волокна, являющиеся, очевидно, центральным продолжением наружных тонких корешков. Следует, однако, заметить, что в задней комиссуре новорожденных все еще содержится значительно более скудное количество волокон, нежели у взрослых. Обстоятельство это может быть объяснено только тем, что, кроме вышеуказанных волокон, в задней комиссуре проходят еще и другие, поздно развивающиеся волокна. Большинство последних, мне кажется, суть не что иное, как комиссуральные волокна, соединяющие между собой клеточные элементы той и другой половины серого вещества.
К ВОПРОСУ О НАРУЖНЫХ СОЧЕТАТЕЛЬНЫХ ВОЛОКНАХ МОЗГОВОЙ КОРЫ
Т. Каес1, описывая в последнее время тонкие волокна мозговой коры, указывает, между прочим, на содержание в последней особого пучка волокон, называемого им на ружным мей-нертовским ассоциационнымслоем. По объяснению автора, в то время как собственно мейнертовские волокна (librae propriae) располагаются в пределах самых наружных границ мозгового вещества (следовательно, под корковым слоем), описываемые им волокна заложены в сером веществе коры в виде прямого дополнения к предыдущим и с непосредственным переходом в них.
Они образуют собой, таким образом, широкие полосы довольно тонких волокон с параллельным расположением по отношению как к мейнертовским волокнам, так и к волокнам промежуточного слоя. Самые наружные пучки этих волокон соответствуют в затылочных и в лобных долях полоске Дженнари или Байярже. Автор высказывает при этом предположение, что будто бы вся кора на всем ее протяжении занята большим, вертикально расположенным по отношению к проекционным волокнам, пучком, который может быть разделен на три отдела: тангенциальный (самый наружный), промежуточный и наружный ассоциацион-ный слой." На особенно удачных препаратах эта поперечная волокнистость коры (Querfaserung) непосредственно переходит от промежуточного слоя в тангенциальный с одной стороны и в наружный ассоциационный слой с другой. Отдельные, по большей части более толстые, косвенно или поперек проходящие волокна встречаются повсюду, но в относительно небольшом числе; в наружном же ассоциационном слое такие волокна встречаются очень редко.
Это описание волокон, проходящих в мозговой коре наперекрест с вертикальными пучками, довольно близко подходит к результатам моих новейших исследований по тому же предмету. Я нахожу даже, что в некоторых областях мозговой коры ассо-циационные или сочетательные пучки, являющиеся как бы допол-
1 К а е s, Die Anwendung der Wolters'schen Methode auf die feinen Fasern der Hirnrin de, Neurol. Zbl., 1891, № 15.
47
нением к fibrae propriae Meynerti, но расположенные внутри мозговой коры, представляются даже более развитыми, чем сами fibrae propriae.
Не лишним считаю здесь заметить, что еще несколько лет тому назад в составленной мною главе о мозговых полушариях, помещенной в «Основаниях микроскопической анатомии», изданных под редакцией М. Лавдовского и Ф. Овсянникова (см. стр. 928), мною указано на существование волокон, проходящих в лучистом слое аммонова рога наперекрест с длинными верхушечными отростками пирамидальных клеток. Как по своему расположению, так и по направлению эти волокна, без сомнения, вполне аналогичны волокнам, описываемым Каесом для других областей мозговой коры.
Я, впрочем, не вполне согласен с Каесом в том отношении, что поперечные волокна мозговой коры принадлежат к числу тонких волокон. По моим исследованиям, в состав наружных ассоциа-ционных волокон, проходящих внутри мозговой коры как в глубоких, так и в более наружных ее отделах, входят как тонкие, так и более толстые волокна, местами же последние преобладают.
В существующих анатомических описаниях мозговой коры (Экснер, Швальбе, Раубер, Тучек и др.) я не встречаю, кроме того, указаний на существование в некоторых областях заднего отдела полушарий особого слоя довольно толстых мякотных волокон, располагающихся довольно тесными рядами при основа-' нии первого слоя (на границе его со вторым). Направление этих волокон (на существование которых мною было указано впервые также в цитированной выше главе о мозговых полушариях и которые уже тогда в отличие от подкорковых fibrae propriae Meynerti были названы мною наружной ассоциационной системо й) представляется приблизительно параллельным поверхности мозговых извилин и обычно соответствует длине последних; поэтому на разрезах, проведенных поперек мозговых извилин, рассматриваемые волокна являются в большинстве случаев поперечно перерезанными. Поразительного развития этот слой достигает в особенности в subiculum cornu Ammonis и в самом аммоновом роге, где он располагается при основании так называемого мелкозернистого слоя, соответствующего первому слою других областей мозговой коры. Здесь этот слой был описан Оберштейнером1 под названием stratum medullare medium и совершенно независимо от него — мною2. Достойно внимания, что в аммоновом роге направление волокон рассматриваемого слоя соответствует длине извилины аммонова рога, вследствие чего на поперечных разрезах они оказываются перерезанными
1 Оберштейнер, Руководство к изучению строения центральной нервной системы, М., 1888, стр. 363.
2 См. рисунок и описание аммоиова рога в составленной мною главе о мозговых полушариях в «Основаниях микроскопической анатомии», под ред. М. Лавдовского и Ф. Овсянникова (стр. 928 и след.).
48
поперек. Следует, впрочем, заметить, что в переходной области между subiculum и cornu Ammonis, т. е. при самом начале завитка последнего, только что указанный слой отсутствует или по крайней мере значительно редуцирован. Зато в этой области можно видеть иногда непосредственный переход волокон рассматриваемого слоя subiculi в так называемую substantia reticularis alba.
Следует заметить, что этот слой, протягиваясь почти через всю ширину аммонова рога, не переходит непосредственно в в fascia dentata в виде такого же слоя, но волокна его, изменив свое направление на косвенное, при завороте аммонова рога в направлении к fascia dentata, переходят широким пучком через упомянутый заворот к основанию fasciae dentatae и, проникая затем через лежащий здесь слой больших пирамидальных клеток, теряются там или между последними, или же вблизи основания только что упомянутого слоя в тончайшем сплетении волоконец, образованных разветвлениями цилиндрических отростков вышеуказанных клеток (в так называемом мелкозернистом слое stratum granulosum Cupferi).
Таким образом, рассматриваемые волокна подходят на довольно близкое расстояние к alveus, но, как я могу убедиться на основании исследования препаратов, окрашенных по способу Экснера и Вейгерта, они не переходят в него, а имеют ближайшее отношение к большим пирамидальным клеткам как самого аммонова рога, так и fasciae dentatae.
Отсюда очевидно, что рассматриваемые нами волокна, подобно волокнам, содержащимся в самом наружном или так называемом тангенциальном слое (resp. substantia reticularis alba аммонова рога), обязаны своим происхождением клеточным элементам нижележащих слоев мозговой коры и представляют собой систему, совершенно аналогичную с простыми сочетательными пучками Мейнерта (librae propriae).
4 В. М. Iiexxcj or;
К ВОПРОСУ О МОЗГОВЫХ ПОЛОСКАХ ПРОДОЛГОВАТОГО МОЗГА
(striae medullares, s. acusticae)
Относительно происхождения striae medullares до сих пор еще не имеется достоверных сведений. Большинство авторов держится того мнения, что они являются продолжением волокон задней ветви слухового нерва. Еще недавно это мнение нашло поддержку со стороны Монакова1, признавшего их частью слухового пути, который по его исследованиям, произведенным с помощью гуд-деновского метода, составляется у кошки из следующих частей: tuberculum acusticum, striae acusticae, перекрест в шве, нижняя (боковая) петля и тыльное мозговое вещество верхней оливы (dorsales Mark der oberen Olive), corp. geniculatum mediale и корковая слуховая сфера.
Также и Багинский2 на основании своих исследований, произведенных по методу Гуддена, ставит striae medullares в соотношение с задним корнем п. acustici, но при этом замечает, что указанное соотношение не должно быть большим, так как атрофия их в отношении атрофии заднего корня, вызванной разрушением улитки, оказывается относительно ничтожной. По Багинскому, несомненно, они представляют собой вторичные пучки, которые стоят в тесной связи с соответствующей верхней оливой, а не перекрещиваются в шве, как думает Монаков.
Следует, однако, заметить, что пучки, которые принимаются авторами за striae medullares s. acusticae у кошки и кролика, не могут считаться аналогичными striae medullares человека, на что было обращено внимание как мною3, так и Вирховым4-
1 Mon akow , Korrespondenzblatt f. Schweizer Aerzte, 1887, № 5; реферат в Neurol. Zbl., 1887, S. 201.
2 Baginsky, Ueber den Ursprung und den zentralen Verlauf d. N. acusticus d. Kaninchens und d. Katze, Virch. Arch., 1890, Bd. 119, S. 91.
3 См. мою работу: «Zur Frage iiber den Ursprung der Hornerven und uber die physiologische Bedeutung des N. vestibularis», Neurol. Zbl., 1887, № 9, а также Вестник психиатрии за тот же год.
4 Virchow R., Ueber die Striae acusticae des Menschen, Verhand-!:mgen d. Physiol. Gesellschaft zu Berlin. Sitzung am 24 Februar 1888.
bO
В первом случае дело идет, собственно, о волокнах, которые-были описываемы также мною в цитированной статье1 и которые действительно образуют собой центральные продолжения волокон задней ветви слухового нерва после прерывания их в переднем ядре (и tub. acusticum). Волокна эти огибают веревчатое тело снаружи и сверху, после чего, углубляясь в мозговое вещество, переходят в дугообразные волокна сетевидной формации и перекрещиваются затем в шве над трапециевидным телом (corpus trapezoideum)2. По Монакову, они образуют затем тыльное мозговое вещество верхней оливы противоположной стороны и, присоединяясь к нижней или боковой петле, вместе с волокнами последней поднимаются к заднему четверохолмию3.
Между тем у человека мы имеем пучки волокон, которые хотя также огибают веревчатое тело снаружи и сверху, но зато протягиваются по дну ромбовидной ямки до средней линии и затем углубляются в шов. Относительно этих истинных striae medulla-res человека еще в 1885 г.4 в работе, опубликованной из лаборатории П. Флексига, я заметил, что они развиваются много позднее корешков слухового нерва и потому, очевидно, не могут быть в прямой связи с ним. Затем в позднейшей своей работе5, в которой, между прочим, мною указано, что упомянутые выше волокна, отожествляемые Монаковым с striae medullares, на самом деле не должны быть смешиваемы с истинными striae medullares, я вновь подтверждаю, что эти последние striae medullares, s. acu-sticae человека в силу их позднего развития не могут служить прямым продолжением корешковых волокон слухового нерва. Эти указания я и теперь считаю безусловно верными, несмотря на то, что со времени опубликования моих работ появились цитированные нами исследования Монакова, повидимому, находящиеся в противоречии с вышеизложенными замечаниями.
К подобным же выводам пришел затем и Онуфрович в работе, опубликованной им в 1886 г.6. Он признает, подобно мне, за непреложный факт, что striae medullares не должны быть рассматриваемы как прямые продолжения слухового нерва; они могут быть или вторичными путями, возникающими из tub. acusticum, или же не имеют с п. acusticus ничего общего. Наконец, не очень
1 См. мою работу в Neurol. Zbl., 1887, № 9, и в Вестнике психиатрии, а а тот же год.
2 Багинский, впрочем, как мы видели, отрицает перекрещивание striae medullares в шве.
8 Высказанное мною предположение (см. Neurol. Zbl., 1887, К» 9,-S. 194) об окончании этих волокон после перекрещивания их в шве в осо--бой группе клеточных элементов верхнего этажа я считаю в настоящее время, после вышеуказанных исследований, мало правдоподобным.
4 'См. мою работу: «Ueber die innere Abteilung des Strickkorpers und des achten Hirnnerven», Neurol. Zbl., 1885, S. 147.
5 См. мою работу в Neurol. Zbl., 1887, S. 194, и в Вестнике психиатрии за тот же год.
6 Onufrowitcz, Arch. . Psychiatrie, Bd. XVI, 3.
4* -?t
Давно проф. Флексиг1 в статье, посвященной выяснению отношения заднего четверохолмия к слуховым нервам, подтверждая вновь, что striae medullares у человека получают мякотную обкладку лишь спустя долгое время после рождения, указывает на возможность их связи с нижним четверохолмием, при этом он замечает, что striae medullares по большей части происходят в raphe до передней периферии продолговатого мозга и переходят затем, как допускалось и Мейнертом, в librae arcuatae externae. По Флексигу, вопреки мнению Багинского, они перекрещиваются с брюшной стороны (вентрально) от corpus trapezoidei, в то время как малая (непостоянная) часть их перекрещивается тотчас под серой почвой желудочка, проникая в боковые части продолговатого мозга.
Исследуя за последнее время ход волокон striae medullares на человеческих мозгах, я убедился, что они на самом деле не имеют ашкакого отношения к волокнам слухового нерва.
В этом нетрудно убедиться, уже сравнивая размеры striae medullares и задней ветви слухового нерва. Известно, что первые представляют собой образования довольно непостоянные по своим размерам, в то время как толщина слухового нерва и задней его ветви, в частности, не подвергается заметным для простого глаза колебаниям. Довольно распространено мнение,что колебания в размерах striae medullares объясняются тем, что часть их нередко скрывается в глубине продолговатого мозга и потому представляется невидимой па поверхности желудка, но в этом я не мог убедиться на своих препаратах. Во всяком случае заслуживает внимания то обстоятельство, что не совсем редко встречаются мозги, в которых striae medullares достигают необычайного развития и по своим размерам не могут быть и сравниваемы с задней ветвью слухового нерва. Для своих исследований я именно и пользовался такими мозгами. Уже при наружном осмотре в этих случаях нередко удается вполне убедиться, что волокна striarum rnedullarum не служат продолжением задней ветви слухового нерва. В боковой своей части striae medullares проходят по верхней и наружной поверхностям веревчатого тела над волокнами задней ветви слухового нерва и затем, располагаясь снаружи от tuberculi acustici переходят непосредственно в белое вещество мозжечка в соседстве с flocculus, что иногда удается видеть простым глазом с необычайной ясностью. Дело в том, что те и другие волокна большей частью удается различить друг от друга уже по внешнему виду при внимательном осмотре соответствующей части продолговатого мозга простым глазом или с помощью лупы. Так, волокна задней ветви слухового нерва представляют собой крайне нежные, в заднем своем отделе не покрытые волокнами striarum rnedullarum, сероватые пучки, которые, обогнув снаружи и сверху веревчатое тело при внутренней его границе, вноеь
1 FlechsigP., Neurol. Zbl., 1890, № 4.
:-2
скрываются внутри ткани продолговатого мозга. Между тем striae medullares в тех, конечно, мозгах, в которых они хорошо» выражены, представляют собой толстые мякотные пучки, проходящие от наружных границ веревчатого тела большей частью вплоть до шва и обычно поражающие своим белым цветом. При этом нередко можно видеть, что на уровне веревчатого тела эти пучки не спускаются по наружной стороне последнего вместе с волокнами задней ветви слухового нерва, а, отклоняясь несколько кпереди* переходят непосредственно в прилежащие области белого вещества мозжечка и здесь почти тотчас же исчезают, углубляясь в мозговую ткань. Делая микроскопические срезы из тех мозгов, где striae достигли крайнего развития, также нетрудно убедиться, что с задней ветвью слухового нерва они не имеют ничего общего, а проходят снаружи от нее и от tub. acustici, возникая из соседней с клочком (flocculus) области.
В направлении кнутри striae medullares, как было уже выше упомянуто, обычно достигают средней линии, затем углубляются в шов. На удачных поперечных срезах можно убедиться с положительностью, что striae medullares пучками проходят внутри шва в брюшном (вентральном) направлении и, перекрестившись в нем, спускаются до передней поверхности продолговатого мозга. Обогнув затем противоположную пирамиду снутри и спереди, частью же проникая тонкими пучочками между ее волокнами, striae medullares, несомненно, переходите librae anteriores противоположной стороны продолговатого мозга. На этом пути их удается легко проследить до входа корешков слухового нерва, где striae быстро прерываются и таким образом ускользают от дальнейшего исследования на поперечных срезах.
Необходимо иметь в виду, что внутри шва striae medullares собственно не образуют перекреста в настоящем смысле этого слова, но, следя за отдельными волокнами и пучками, можно все же убедиться, что они переходят внутри шва на другую сторону. Достойно внимания также то обстоятельство, что шов на всем своем протяжении в месте прохождения striatum medullaruni занят серым веществом, внутри которого собственно и проходят striae. Имеет ли это серое вещество, стоящее в непрерывной связи с серым веществом мозга, непосредственное отношение к волокнам striae, я решить не берусь, но несомненно, что так называемое ядро пирамиды (п. arciformis) служит местом прерывания части волокон striae. По крайней мере на препаратах можно видеть, что в упомянутое ядро вступает часть волокон последних и рассыпается в нем в тончайшую сеть, тогда как и с другой стороны ядра выходят из него волокна, присоединяющиеся к пучку striae. Впрочем, большинство волокон последних минует ядро пирамиды, переходя непосредственно в librae arcuatae anteriores. Несомненно, что ни к п. funiculi teretis, ни к п. vagus и glossopharyngeus, ни к п. trigeminus striae, medullares, вопреки мнению некоторых авторов, не имеют решительно никакого отношения. Я сомне-
53
ваюсь также, чтобы они имели ближайшее отношение к заднему двухолмию. Более всего правдоподобным мне кажется предположение, что striae medullares служат для связи между собой основных областей мозжечка. С этим вместе было бы нетрудно объяснить как непостоянство в относительном развитии этих пучков, так и нередко встречающиеся своеобразные отступления в ходе их.
Известно, например, что в некоторых случаях часть striae medullares по достижении шва непосредственно переходит на другую сторону. В иных случаях striae medullares в виде более или менее компактных пучков идут от срединной бороздки в косвенном направлении кпереди и кнаружи, погружаясь в среднюю ножку мозжечка на довольно значительном расстоянии от тыльной поверхности веревчатого тела (так называемый Klangstab немцев, conductor sonorus Бергмана, непостоянный корень тройничного нерва, по Штиллингу).
Выше было упомянуто, что striae возникают из белого вещества мозжечка, соседнего с клочком (flocculus). He следует, однако, смешивать волокна striarum с ножкой клочка. Последняя представляет собой вполне самостоятельный и хорошо обособленный пучок, облагающийся миэлином в относительно раннем возрасте, во всяком случае за несколько недель до рождения (у зародышей величиной за 38—40 см он представляется уже мякот-ным), в то время как волокна соседних частей мозжечковых полушарий и striae medullares, развивающиеся лишь после рождения, представляются еще совершенно безмякотными. Благодаря этому на окрашенных по Вейгерту или Палю разрезах, приготовленных из мозгов незрелых новорожденных, указанный пучок волокон всегда выступает с поразительной ясностью, причем нетрудно убедиться, что он образуется из волокон, берущих начало в задне-основной области клочка (flocculus). Выходя из коры извилинок последнего, волокна этого пучка, как я мог проследить на целом ряде разрезов, приготовленных частью мною самим, частью студентом-медиком Купревичем, направляются первоначально прямо кнутри по основной поверхности клочка, а затем поднимаются по краю мозжечкового полушария, облегающему веревчатое тело при вступлении последнего в мозжечок, и таким образом достигают бокового края дна IV желудочка. Отсюда большинство волокон описываемого пучка поднимается по кровле желудочка в мозжечок, где их удается проследить вплоть до уровня corpus dentati cerebelli.
В заключение скажу несколько слов по поводу неоднократно затрагиваемого в последнее время вопроса о первенстве открытия некоторых фактов, имеющих отношение к центральным окончаниям слухового нерва.
Еще в 1885 г. в № 7 «Neurologisches Zentralblatt» мною была опубликована вкратце работа из лаборатории профессора Флек-сига,.существенные результаты которой сводятся к следующему:
54
1. Слуховой нерв распадается по времени развития на д в а хорошо различимых отдела: а) несколько ранее развивающийся отдел этого нерва соответствует в существенном «переднему» корню слухового нерва, обнимает собой все входящие в центральный орган кнутри corpus restiforme волокна и происходит из п. vestibularis, вследствие чего он и назван был мною корнем п. vestibularis; б) познее развивающийся отдел слухового нерва, соответствующий «заднем у» корню авторой, содержит все расположенные кнаружи от corpus restiforme волокна и выходит из п. cochlearis, вследствие чего он был назван мною корнем п. vestibularis.
2. Ни один из этих корней не имеет прямой связи с мозжечком.
3. Корень п. vestibularis частью оканчивается в серой массе, которая расположена в боковой части IV желудочка дор-зально от ядра Дейтерса, частью же спускается вдоль последнего к продолговатому мозгу. Корень п. cochlearis оканчивается большей частью в переднем ядре слухового нерва, из которого выходит corpus trapezoideum (Флексиг).
4. Striae medullares развиваются много позднее обоих корней слухового нерва и потому, очевидно, не могут стоять в прямой связи с последним.
Две недели ранее того в моей работе под заглавием «Zur Ana-tomie der Schenkel des Kleinhirns, insbesondere der Bruckenarme {Neurol. Zbl., 1885, № 6, S. 122), приведено указание следующего рода: «Corpus trapezoideum берет, в основном, начало в переднем ядре слухового нерва и представляет собой центральный проводящий путь слухового нерва, иначе говоря, VIII пары черепных нервов» (Флексиг).
Далее в моей работе «О петельном слое» («Ueber die Schlei-fenschicht»); сообщенной проф. П. Флексигом в заседании Королевского саксонского ученого общества 4/V 1885 г.1 указывается впервые, что исследование мозгов, принадлежащих зародышам 26—30 см, принуждает к принятию, что ядро нижнего четверохолмия через боковую петлю соединяется с верхней оливой и corpus trapezoideum (а через то с восьмым нервом)3. Затем в работе «О соединении верхних олив и их вероятной физиологической роли», опубликованной в том же году на русском языке* и реферированной в № 21 «Neurologisches Zentralblatt» за 1885 г., я выяснил, между прочим, на основании исследования зародышевых мозгов, что нижняя или боковая петля образует собой пучок волокон, который берет начало главным образом из соответствующей, отчасти же из противоположной верхней оливы и устанавливает, таким образом, связь между этими сб-
1 См. реферат в Neurol. Zbl., 1885, № 3, S. 353.
2 Все подчеркнутое в вышеприведенных цитатах было подчеркнуто и в тексте работ, из которых заимствованы эти выдержки.
3 Врач, 1885, № 32; реф. Neurol. Zbl., 1885, № 21.
оо
разованиями и задним четверохолмием. С другой стороны, там же указана мною связь между верхними оливами и передним ядром слухового нерва при посредстве поперечных волокон corpus trapezoidei. Наконец, в статье «О нервных путях спинного и головного мозга», помещенной в «Основаниях к изученю микроскопической анатомии человека и животных» (под ред. М. Лавдов-ского и Ф. Овсянникова), на стр. 949 я буквально говорю следующее: «на разрезах из зародышевых мозгов удается показать связь верхних олив с наружным, ядром слухового нерва (передним Мейнерта), принимающим в себя задний корень последнего, и с ядром отводящего нерва. Волокна, соединяющие верхние оливы с наружным слуховым ядром (18, фиг. III и VI), выходят из последнего прямо кнутри наперекрест с передним корнем слухового нерва; они только частью рассеиваются в соответствующей верхней оливе, большинство же переходит в поперечные волокна трапециевидного тела и, перекрещиваясь затем в raphe, поднимается в нижней или боковой г петле к заднему четверохолмию». Нелишним считаю заметить здесь, что на приложенном к статье схематическом рисунке вышеозначенный путь, соединяющий слуховое ядро с задним четверохолмием, и дальнейшие его продолжения в коре-обозначены вполне соответственно содержанию текста.
До опубликования первой из этих работ появилось в печати лишь краткое сообщение Фореля 3, опередившее опубликование моей работы на один месяц. Но в этом сообщении Фореля я нахожу лишь то общее с данными моей работы, что слуховой нерв имеет два корня, которые, как известно, различались уже и прежними авторами, и что задний корень большей частью своих волокон оканчивается в переднем ядре слухового нерва. Об отношении же переднего корня к преддверной части и заднего корня к улитковой части слухового нерва в этой работе не упоминается ни полсловом.
Таким образом, очевидно, что, за указанным исключением, вышеприведенные факты были заявлены мною и Флексигом впервые и притом заявлены вполне категорично в столь распространенном реферирующем печатном органе, как «Neurologisches Zentralblatt». Ввиду этого кажутся в высшей степени странными сделанные в свое время со стороны Онуфровича заявления, клонящиеся к умалению значения наших исследований и к отрицанию первенства (Prioritat) сделанного нами открытия, что улитковая часть слухового нерва имеет своим центральным продолжением задний корешок, а преддверная часть — передний корешок слухового нерва. Эти заявления в свое время встретили совершенно справедливый протест со стороны проф. Флексига,
1 Тождественные указания можно найти и в моей статье «Le cerveau de l'homme dens ses rapports et connexions intimes», Arch, slaves de biolo-gie, 1887.
2 Cm. Neurol. Zbl., 1885, № 5.
56
высказанный им в статье «Zur Lehre vom zentralen Ve/lauf der Sinnesnerven» г.
He очень давно, однако же, Багинский в одной из своих работ2 снова прибегнул к подобному же приему, как и Онуфрович, в отношении доказанной мной и Флексигом связи слухового нерва с задним четверохолмием. Несмотря на то, что в вышеци-тированных наших работах, как мы видели, связь переднего ядра слухового нерва с задним четверохолмием при посредстве волокон corpus trapezoidei и задней или боковой петли многократно и категорически утверждалась, Багинский силится доказать, что мы высказали лишь предположение или гипотезу, а что доказательство связи слухового нерва с задним четверохолмием принадлежит ему: «Охотно признаю, что эта гипотеза принадлежит Бехтереву и Флексигу; однако доказательство она нашла лишь в моих экспериментальных исследованиях».
В чем же причина того, что Багинский наши заключения считает лишь простым предположением, а не доказанным фактом? Виной тому оказываются скобки, в которые заключены шесть слов в одной из цитированных выше работ.
Этому, конечно, не всякий поверит, а потому считаю не лишним привести слова Багинского целиком: «Будучи далек от того, чтобы вступать в спор о приоритете, я все же позволю себе заметить, что в отчете о заседании Саксонского королевского научного общества, на который ссылается Флексиг и который в момент первичного сообщения о моей работе был мне неизвестен3, попав мне в руки лишь позднее благодаря любезности Флексига, слова «и тем самым с VIII парой черепных нерЕов» взяты в кавычки, опущенные лишь после опубликования моей работы (sic!). Таким образом, совершенно очевидно, что Флексиг (и я!) в 1885 году не наблюдал связи между слуховым нервом и задним четверохолмием. Кроме того, выражение „заставляет предположить", как мне думается, приведено лишь для того, чтобы дать повод для сомнения».
Всякие пояснения к этой выдержке я нахожу совершенно излишними, так как и без того ясно, что ближайшим поводом к такого рода заявлениям может служить лишь совершенно неуместная и неоправдываемая фактами погоня за первенством.
1 Neurol. Zbl., 1886, № 23.
2 В a g i η s k у , Ueber den Ursprung und den zentralen Verlauf des-N. acusticus des Kaninchens und der K.atze,Virch. Arch., Bd. 119, S. 81 u.f.f.
3 Неизвестной Багинскому, собственно, осталась не одна цитируемая им моя работа, но и три других моих сообщения, касающиеся того же предмета (см. выше).
ОБ «ОЛИВЧАТОМ» ПУЧКЕ ШЕЙНОЙ ЧАСТИ СПИННОГО МОЗГА
В начале марта настоящего года я получил от д-ра Гельвега из Дании любезное письмо вместе с отдельным оттиском его статьи «Studien iiber den zentralen Verlauf der vasomotorischen Nervenbahnen», помещенной в Archiv fur Psichiatrie, Bd. XIX, Hft. I. В этом письме д-р Гельвег выражает сожаление, что он своевременно не имел возможности узнать о содержании моей работы,в которой мною впервые (1885) описан «центральный пучок покрышки», выходящий из нижних олив в черепном направлении. Из отдельного оттиска статьи д-ра Гельвега представляется очевидным, что он описывает в своей работе, появившейся много позднее моей, тот же самый пучок, который был уже описан мною под названием центрального пучка покрышки 1. Мне приходится в свою очередь пожалеть, что содержание работы д-ра Гельвега мне оставалось неизвестным до позднейшего времени, но пожалеть по совершенно иному поводу. Дело в том, что в своей статье под вышеприведенным заглавием, мало говорящим об анатомическом содержании работы, автор описывает спинную часть, повидимому, той же системы волокон, которая в мозговом стволе образует собой описанный мною центральный пучок покрышки и которую автор называет треугольным пучком (dreikantige Bahn).
Пучок этот мне был известен уже давно, еще со времени моих занятий в лаборатории профессора Флексига. В заведуемои мною психофизиологической лаборатории Казанского университета я его неоднократно демонстрировал занимающимся врачам и, между прочим, имел в виду совместно с занимавшимся в моей лаборатории д-ром Останковым всесторонне обследовать этот пучок на мозгах новорожденных. К сожалению, большое количество занятий, с одной стороны, а с другой — мое служебное перемещение из Казани в С.-Петербург помешало осуществлению этого намерения.
Все это я говорю не с тем, чтобы заявить первенство открытия вышеуказанного пучка, а для того, чтобы подтвердить дей-
1 В е с h t е г е w W., Ueber eine bisher unbekannte Verbindung der grossen Oliven mit dem Grosshirn, Neurol. Zbl., 1885, № 9.
-58
ствигельяое его существование, так как полагаю, что нет лучшей гарантии в точности какого-либо открытия, если оно заявляется с двух различных сторон и притом совершенно независимо одна от другой.
В отличие от Гельвега, исследования которого главным образом произведены на мозгах душевнобольных, мои исследования, касающиеся вышеуказанного пучка, производились главным образом по методу развития. Несмотря на это различие методов, я должен заметить, что данные моего исследования в существенных чертах вполне согласны с результатами исследований Гельвега и в этом я снова вижу гарантию в точности наших результатов.
Рис. 1.
Рис. 2.
Пучок, о котором идеть речь, как я убедился, принадлежит •к поздно развивающимся системам спинного мозга, так как он облагается мякотью даже позже пирамидного пучка и во всяком случае развитие его заканчивается лишь по рождении плода на свет.
Нечего и говорить, что этот пучок развивается много позднее соседнего с ним передне-наружного пучка, описанного Говер-сом и независимо от него мною.
Благодаря этому, рассматриваемый пучок хорошо выделяется на мозгах новорожденных, у которых в спинном мозгу, кроме вышеуказанного пучка, представляются безмякотными лишь одни пирамидные пучки. Исследование, производимое на таких мозгах, убеждает нас, что рассматриваемый пучок принадлежит главным образом шейной части спинного мозга и отчасти нижнему отделу продолговатого мозга, так как впервые он обнаруживается на поперечных срезах в виде небольшого мениска между передними корешками на уровне верхнего отдела шейного утолщения и затем исчезает при нижнем конце больших олив (см. рис. 1 и 2, /о).
В своей нижней части пучок этот располагается на границе между передним и боковым столбами, в области выхода передних корешков, но затем в направлении кверху, быстро пополняясь своими волокнами, он продвигается все более и более вперед и
59
при переходе в продолговатый мозг достигает даже до области' передних пирамидных пучков.
Что касается tro внешней формы на поперечных срезах, то название «треугольный», даваемое этому пучку Гельвегом, может быть применимо только к незначительному отделу этого пучка, заложенному в средней части шейной области спинного мозга; ниже, при своем исходе, пучок этот утрачивает треугольную форму, а имеет вид скорее небольшого мениска (см. рис. 1, /о); точно так же и при переходе в продолговаты]'! мозг он совершенно утрачивает свою треугольную форму.
Как упомянуто, при переходе спинного в продолговатый мозг рассматриваемый пучок постепенно смещается все более и более вперед; в нижней части продолговатого мозга он уже почти соприкасается с пирамидой, а затем, при появлении большой "оливы, быстро исчезает, скрываясь в передне-наружном отделе последней.
Ввиду того что наш пучок быстро нарастает по направлению снизу вверх, он, несмотря на свое относительно короткое протяжение, при переходе в продолговатый мозг занимает довольно значительные размеры.
Его волокна, как справедливо замечает Гельвег, принадлежат в большинстве случаев к числу очень тонких волокон, но я сомневаюсь пока, чтобы тонкие волокна, рассеянные в окружающей области основного пучка, в так .называемой смешанной системе Флексига (gemischtes System) и в соседстве с передним рогом, принадлежали, как думает Гельвег, к одной системе с рассматриваемым пучком. Дело в том, что у новорожденных в возрасте первого месяца этот пучок представляется еще безмякотным, волокна же основного пучка уже почти сплошь обложены мякотью.
Благодаря ближайшему отношению рассматриваемого пучка к большим или нижним оливам, я нахожу правильным называть его оливчатым п у ч к о м шейной части спинного мозга.
Так как с другой стороны нижних олив из последних выходит в черепном направлении описанный мною впервые центральный пучок покрышки, то очевидно, что оливчатый пучок шейной части спинного мозга вместе с последним принадлежит к системе волокон, прерываемой на своем пути нижними оливами, от которых, кроме того, как известно, отходят многочисленные волоки.?, направляющиеся в противоположное веревчатое тело и затем в мозжечок.
В заключение необходимо упомянуть, что, благодаря относительно позднему развитию, рассматриваемый пучок на мозгах новорожденных в высшей степени легко выделяется от рано развивающегося передне-наружного пучка, описанного впервые Говерсом и мною х. Должно, впрочем, заметить, что Говерс не
1 В е с h t е г е w W. M., Neuro!. Zbl., 1885, S. 155, u. Go w e r s. ibid, 188(i, S. 97—98 v. 150.
60
совсем правильно в одном из своих сообщений по поводу передне-наружного пучка замечает, что пучок этот протягивается по периферии спинного мозга вплоть до переднего пирамидного пучка *; это не совсем согласно с моими исследованиями по методу развития, произведенными, как я указывал ранее2, совершенно независимо от наблюдений Говерса: при исследовании зародышевых мозгов мне не удалось проследить передне-наружный пучок за областью передних корешков. Говерс объясняет разногласие особенной тонкостью пучка по ту сторону корешков3 ; но несомненно, что к верхней части шейной области это объяснение неприменимо, ввиду того, что здесь периферия переднего столба, как видно из предыдущего, занята позднее развивающимся олив-чатым пучком.
Neurol. Zbl., 1886, S. 150.
См. мои «Проводящие пучки мозга», отдельное издание.
Neurol. Zbl., 1886, S. 98.
ОБ ЯДРАХ ГЛАЗОДВИГАТЕЛЬНЫХ НЕРВОВ
(NN.OCULOMOTORII, ABDUCENTIS И TROCHLEAR IS)
И О СВЯЗИ ИХ ДРУГ С ДРУГОМ
(с 11 рисунками в тексте)
Знакомясь с анатомической литературой, посвященной ядрам трех глазных нервов, мы встречаемся со многими, далеко еще не выясненными вопросами, в силу чего дальнейшие исследования
в отношении вышеуказанного предмета на мой взгляд представляются вообще крайне желательными.
Исследовав большое число младенческих и зародышевых мозгов человека на последовательном ряде срезов, я убедился, что к п. oculomotorius принадлежат собственно четыре ядра — два больших, из которых одно парное и одно непарное, и два меньшей величины, или добавочные, ядра — оба парные.
К первым принадлежит прежде всего главное ядро (тыльное ядро авторов), располагающееся под передним двухолмием по обе стороны срединной линии и имеющее на поперечных срезах полулунную форму (рис. 1). Оно является наибольшим из всех вообще ядер глазодвительного нерва и содержит средней величины многоотростчатые нервные клетки,заложенные в густом сплетении волоконец. С наружно-брюшной стороны к этому ядру непосредственно прилежат волокна заднего продольного пучка, причем часть серого вещества, принадлежащего ядру, даже отделяется от его массы и располагается снаружи и с брюшной стороны заднего продольного пучка; благодаря этому на поперечных срезах получается впечатление, как будто задний про-
Рис. 1.
nd—заднее или главное ядро; оси/—корешки п. oculomotoril; /рг—волокна глубокого вещества четверохолмия, проникающие к срединному ядру; fls—поперечно перерезанные волокна заднего продольного пучка. Зародышевый мозг, окраска по Weigert'y.
о2
дольный пучок проникает через наружно-брюшные отделы ядра. Вместе с тем оба ядра в своей средней части соприкасаются и до> известной степени даже сливаются своими внутренними брюшными краями (рис. 2, nd)
Рис. 2. Срез на уровне ядер п. oculomotorii.
nd—боковое и.*?и заднее ядро п. oculomotorii; пат—срединное ядро; пай — добавочные ядра п. oculomotorii; fp—пучок глубокого мозгового вещества четверохолмия* подходящий к ядрам п. oculomotorii.
Рис. 3. Срез на уровне ядер п. oculomotorii.
nd ~~ заднее или главное ядро; пт — срединное ядро; па—добавочное ядро, располагающееся сзади и отчасти снаружи от главного ядра п. oculomotorii; fpr — пучок глубокого мозгового вещества четверохолмия, направляющийся частью к добавочному ядру.
На уровне средней и передней трети главных ядер обнаруживается еще особое, содержащее внутри себя значительные по размерам нервные клетки, непарное срединное ядро, располагающееся по срединной линии между обоими предыдущими ядрами
63-
Рис. 4. Срез на уровне ядер п. oculo-motorii.
nd-бохоъое или заднее ядро; пт—срединное
или внутреннее ядро; fp —пучок глубокого
мозгового вещества четверохолмия, подходя-
^щий к ядрам п. oculomotorii.
и представляющееся много меньшей величины по сравнению с ними (рис. 3, пт и рис. 4, пт).
Далее с тыльной, отчасти же с тыльно-наружной стороны от главных ядер располагается небольшое парное образование, по-видимому, не вполне обособленное от последних (рис. 2, пай и рис. ?, па). Наконец, на уровне передних отделов главного ядра и впереди от срединного ядра по обеим сторонам срединной линии располагается небольшой величины парное ядро закругленной формы (рис. 5, па').
Оба последних ядра, которые я назывлюдобавочными, содержат более нежные и более поздно развивающиеся корешковые волокна, а потому и обнаруживаются лучше всего на мозгах более позднего возраста. Следует еще упомянуть, что наиболее тыльная часть переднего отдела главного_ядра местами как бы обособляется от остальной части главного ядра, не представляя собой, впрочем, самостоятельного образования (рис. 4).
На уровне переднего отдела главного ядра в тыльно-наружном направлении и несколько поодаль от него мы встречаем еще одно умеренной величины образование, располагающееся при боковом крае центрального серого вещества (рис. 5 и 6, пер).
Ядро это, принятое Л. О. Даркшевичем за верхнее ядро п. oculomotorii, в действительности не имеет никакого отношения к корешкам этого нерва, в чем я вполне схожусь с сделанными в этом направлении указаниями Келликера и новейшими исследованиями Берпгеймера. Вышеуказанное
Рис. 5. Часть среза из новорожденного на уровне красных ядер непосредственно впереди от главного ядра п. oculomotorii.
«г—красное ядро; пер—ядро задней спайки; па' — передне-внутреннее добавочное ядро глазодвигательного нерва; fls—окончание заднего продольного пучка.
64
ядро принадлежит, несомненно, к брюшному отделу задней спайки, который, как я показал впервые, развивается в очень раннем возрасте, представляясь мякотным уже у зародышей около 28 см, следовательно, много ранее тыльного отдела задней спайки1.
Заметим здесь, что некоторыми авторами, как Вестфалем и Зимерлингом2, под названием nucl. ventralis post. nucl. oculo-motorii опсывалось содержащее большие многоугольные клетки ядро, расположенное несколько ниже или позади главного ядра и лежащее частью с тылу, частью внутри заднего продольного пучка. Ядро это, как мы впоследствии увидим, в действительности принадлежит боковому нерву и не имеет ничего общего с п. oculomotorius.
Большая часть корешковых волокон п. oculo-motorii возникает из ядер той же стороны и направляется частью в брюшном, частью в наружно-брюшном направлении к месту выхода из мозга; меньшая часть волокон п. oculomo-torii подвергается перекрещиванию между тыльными ядрами и, следовательно, зозникает из ядер противолежащей стороны (рис.7).
К перекрещивающимся волокнам принадлежат собственно задние корешковые волокна п. oculomo-torii, на что в последнее время обратил внимание Бернгеймер; к неперекрещивающимся принадлежат все располагающиеся более кпереди. Первые берут начало в задней половине главного ядра, вторые — в передней его половине, в срединном ядре и в обоих добавочных ядрах.
Задний продольный пучок, несомненно, скрывается частью своих волокон в ядрах п. oculomotorii, преимущественно в глав-
1 См. Даркшевич, Ueber die hintere Komrnissur, Neurol. Zbl., 1884.
2 Arch. f. Psych., Bd. XXIJ.
Рис. 6. Срез из мозга несколько недельного младенца на уровне красного ядра pulv — pulvinar thalami; stz — str. zonale.
fbp—волокна задней ручки; //g—волокна петлевого слоя, начинающие рассеиваться при переходе в thalamus; lsp—волокна петлевого слоя, перешедшие в нижний этаж мозговой ножки; fsn—волокна subst. nlgrae; р—пирамидный пучок; «—нижний отдел corp. subtha-lamici; sn—subst.nigra; nr—n. ruber; fc — волокна центрального пучка покрышки; пер— ядро задней спайки; ср — задняя спайка; Is—пучок, выходящий из ядра заднего двухолмия к зрительному бугру. Окраска по Weigert'y.
О В. В. Бехтерев
65
ndin
ndlll
ном и, может быть, в срединном ядре. Переход волокон заднего продольного пучка в добавочные ядра для меня представляется пока вопросом.
Остальная часть заднего продольного пучка, не скрывающаяся в ядрах п. oculomotorii, на уровне передних отделов главного ядра, начинает постепенно отодвигаться в тыльно-на-ружном направлении по краю центрального серого вещества и вступает в вышеуказанное ядро, принадлежащее брюшному отделу задней спайки. Таким образом, часть заднего продольного пучка является, несомненно, продолжением брюшного отдела задней спайки, другая же часть имеет ближайшее отношение к ядрам п. oculomotorii.
Рис. 7. Препарат из 7-месячного человеческого плода.
///-корешки п. oculomotorii;' nd ///—заднее ядро п. oculomotorii; cr — перекрест корешковых волокон п. oculomotorii между задними ядрами. Окраска по Weigert'y.
Рис. 8. Срез из мозга 6-месячиого человеческого плода.
cbp — ядро заднего двухолмия; срЬ—corp. parabigeminium или nucl. iemnisci lateralis; lc—наружный отдел главной петли, содержащий волокна из ядер клиновидных пучков; flp — задний продольны.й пучок; IV — корешки п. abducentis; nIV— ядро п. abducentis; fniv — волокна, выходящие из ядра п. abducentis в область заднего продольного пучка.
С другой стороны, к ядрам п. oculomotorii подходят волокна глубокого отдела переднего двухолмия, веерообразно расходящиеся в области серого вещества последнего. Большая часть этих волокон образует с брюшной стороны от центрального серого
60
вещества так называемый фонтановидный перекрест и спускается затем в нисходящем направлении во внутренних отделах сете-видного образования к спинному мозгу. Меньшая же часть, несомненно, вступает в ядра п. oculomotorii. Таким образом, часть волокон заднего продольного пучка, возникающего из ядер п. oculomotorii, является, по крайней мере частью, продолжением волокон глубокого отдела переднего двухолмия.
Что касается ядер блокового нерва, то в этом отношении до позднейшего времени существовали не незначительные разноречия в литературе; так, например, за главное ядро п. trochlearis неко торыми авторами и, между прочим, Вестфалем и Зимер-лингом признавалось ядро, содержащее довольно большие закругленные клеточные элементы и располагающееся на уровне заднего двухолмия внутри центрального серого канала в тыльном и отчасти в тыльно-наружном направлении от заднего продольного пучка. С другой стороны, кругловатая группа мелких клеточных элементов, прилежащая к хвостовой части выше описанного ядра, была описана Вестфалем под названием заднего мел ко клеточного ядра п. trochlearis. В последнее время против принадлежности того и другого ядра к trochlearis высказался Кауш \ который за истинное ядро п. trochlearis признает ядро расположенное на тех же уровнях внутри заднего продольного пучка и признаваемое некоторыми авторами за п. centralis posterior п. oculomotorii (см. выше). Эти исследования Кауша, однако, не остались без возражения, в силу чего, мне кажется в этом отношении могут иметь значение указания, сделанные на основании эмбриологического метода.
Вопрос о принадлежности вышеупомянутого ядра к п. trochlearis может быть с точностью выяснен на зародышевых мозгах в силу того обстоятельства, что корешковые волокна, как известно,
Рис. 9. Косвенный разрез через мозговой ствол сверху и сзади вперед и вниз из мозга несколько недельного младенца.
nse—область верхняя внутреннего центрального ядра; flp—задний продольный пучок; !с—центральный пучок покрышки; IV—корешки блокового нерва после перекреста над aq. Sylvii; II—боковая петля; /—главная петля; lsp — волокна петлевого слоя, переходящие в мозговую ножку; р—пира мидиый пучок; /sp'— рассеянные пучки тонких волокон внутри петлевого слоя; сгса — перекрест волокон передней ножки мозжечка. Окраска по Weigert'y.
К a u s с h, Neurol. Zbl., 1894, № 14.
6'.'
развиваются очень рано, волокна же окружающие центральное серое вещество, исключая задний продольный пучок, развиваются в более позднем возрасте. Пользуясь подходящим материалом из человеческих зародышей, я имел полную возможность убедиться в принадлежности вышеуказанного ядра, заложенного внутри заднего продольного пучка к корешкам п. troch-learis, которые могут быть непосредственно прослежены в этом ядре (рис. 8, nIV).
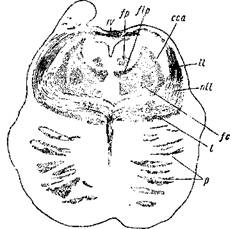
Рис. 10. Срез позади четверохолмия из мозга несколько недельного младенца.
IV — корешки блокового нерва, перекрещивающиеся над aq. Sylvii; fp — область заднего пучка Schutz'a; flp — задний продольный пучок; сса — передняя ножка мозжечка; II—боковая петля; nil—ядро боковой петли; fc—центральный пучок покрышки; /—главная петля; р—пирамидный пучок.
Ввиду того, что некоторые авторы высказывали сомнение в полном перекресте волокон блокового нерва позади заднего двухолмия, заметим здесь, что метод развития в выяснении этого вопроса оказывает незаменимые услуги и ставит вне сомнения существование полного перекреста волокон блокового нерва.
Ход этого нерва на всем протяжении внутри мозга крайне легко проследить на зародышевых мозгах, где его волокна представляются мякотными еще в том возрасте, когда соседние части мозга или вовсе не содержат мякотных волокон, или содержат их лишь в малом количестве. На срезах из таких мозгов можно убедиться, что волокна блокового нерва образуют полный пере-кргст внутри переднего паруса, не перекрещенных же волокон не существует вовсе (рис. 9 и 10, IV).
После перекреста волокна блокового нерва проходят на некотором протяжении в виде круглых пучков снутри от нисходя-
68
щего корешка тройничного нерва, после чего, отклоняясь дугообразно внутрь и в брюшном направлении, они достигают ядра своей стороны, о котором мы упомянули выше (рис. 8, nIV). Что касается ядер п. abducentis, располагающихся в тыльных частях моста под местом загиба корешка личного нерва (рис. И, riVI), то на зародышевых мозгах можно убедиться, что они посылают от себя волокна лишь в корешки соответствующей стороны, следовательно, не существует предполагаемого некоторыми ав-
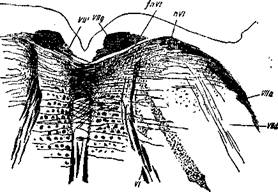
Рис. 11. Разрез на уровне ядра п. abducentis из человеческого плода 33 см длины.
'Vila—восходящая часть корешка facialis; Vlld — нисходящая часть корешка facialis; nVI — ядро п. abducentis; fnVI—волокна, выходящие из ядра п. abducentis к заднему продольному пучку противолежащей стороны; V//g—колено корешка п. facialis; VII'—волокна п. facialis, направляющиеся к противоположной стороне; VI — корешки п. abducentis. Окраска no Weigert'y-
торами неполного перекрещивания корешковых волокон отводящего нерва, что вполне согласно с новейшими исследованиями Бергмана, произведенными по методу перерождения1.
Относительно взаимной связи ядер п. abducentis и п. oculomotorii, как известно, до сих пор остается еще много невыясненного. Правда, уже Дюваль и Лаборд указали на существование волокон, проходящих под дном IV желудочка от ядра п. abducentis одной стороны к ядру п. oculomotorii другой стороны. Однако эти указания не могли быть подтверждены при позднейших исследованиях, а потому в виду доказанного перекрещивания части корешковых волокон п. oculomotorii некоторые авторы принимали, что между тем и другим ядром существует прямая связь при посредстве волокон, проходящих в заднем продольном пучке.
1 Bergmann, Jahrb. f. Psych., Bd. XI.
На основании своих исследований зародышевых мозгов и мозгов новорожденных я убедился, однако, что из каждого ядра отводящего нерва, действительно, отходят в довольно значительном числе волокна внутрь к заднему продольному пучку, который они проникают насквозь и затем, перекрестившись во шве, достигают заднего продольного пучка другой стороны (рис. 11, fnVI). Более чем вероятно, что эти волокна именно и служат для связи ядра отводящего нерва одной стороны с ядром п. oculomotorii другой стороны.
Равным образом на основании эмбриологического материала нетрудно убедиться, что и от ядра п. trochlearis направляется небольшой пучок волокон в область заднего продольного пучка своей стороны; пучок этот, подобно предыдущим волокнам, очевидно, также служит для сочетательной деятельности глазных яблок.
Наконец, и между ядрами п. oculomotorii обеих сторон имеется сочетательная система волокон, которая, без сомнения, также имеет известное значение при совместной деятельности глазных яблок.
05 ОСОБОМ ВНУТРЕННЕМ ПУЧКЕ БОКОВЫХ СТОЛБОВ
Как известно, часть белого вещества боковых столбов, расположенная на границе с серым веществом, была выделена Флек-сигом под названием пограничного слоя. Последний, по Флек-сигу, в верхней шейной области располагается кнаружи от переднего рога, простираясь от передней его группы до заднего конца рога. В шейном утолщении он занимает угол между передним и задним рогом. В грудной области его передняя часть проходит снова вперед и образует пучок, располагающийся снаружи переднего рога; но он представляется здесь тоньше, нежели в верхней шейной области. В поясничном же утолщении отношения этого пучка еще недостаточно выяснены. Что касается состава волокон пограничного слоя, то в нем предполагались частью волокна короткого протяжения, частью волокна из кларковых столбов, частью, наконец, волокна из задних корешков. Не подлежит, впрочем, сомнению, что волокна задних корешков не входят в состав пограничного слоя, так как перерезка задних корешков не влечет за собой перерождения волокон пограничного слоя.С другой стороны, и волокна из столбов Кларка лишь пересекают область пограничного слоя на своем пути к мозжечковому пучку, но не входят в состав продольных волокон.
Уже много лет тому назад, еще во время заведывания мною психофизиологической лабораторией Казанского университета, на основании исследований по методу развития мне удалось выделить в вышеуказанном пограничном слое один пучок, располагающийся в более задних отделах этого слоя, который я назвал внутренним пучком боковых столбов. Об этом пучке я говорю хотя и сжато, но вполне определенно уже в первом издании своих «Проводящих путей», вышедшем в 1893 г. на русском языке и в 1894 г. в переводе на немецкий язык. Равным образом об этом пучке я говорю теми же словами и во втором издании «Проводящих путей», вышедшем на русском языке в 1896 г. 1.
1 См. Проводящие пути спинного и головного мозга. Руководство к изучению внутр~"иих связей мозга, изд. Риккера, 1896.
71
Из этого издания я заимствую здесь следующие строки (см. стр. 99 «Проводящих путей мозга»).
«Основной пучок кзади распространяется частью также и на область, известную под названием пограничного слоя. В последнем, однако, следует различать особый пучок, который в силу его положения я называю внутренним пучком. Волокна этого пучка развиваются позднее волокон основного пучка, но ранее волокон других соседних пучков, благодаря чему он легко может быть выделен на зародышевых мозгах соответствующего возраста (рис. 48 и 49)»1.
«Рассматриваемый пучок с ясностью обнаруживается уже в грудной области спинного мозга, где он располагается непо средственно снаружи от серого вещества между боковым и задним рогом, занимая собою, таким образом, часть пространства, заключающегося между пирамидным пучком и серым веществом спинного мозга, далее же кверху пучок этот продвигается более кпереди, располагаясь все время вблизи серого вещества».
Что касается происхождения волокон внутреннего пучка, то я высказался в том же издании следующим образом:
«Надо думать, что он имеет отношение к клеточным элементам, располагающимся в более задних областях серого вещества, и состоит из коротких волокон, так как перерождение его у человека до сих пор еще не наблюдалось. Впрочем, при перерезке спинного мозга у кролика мне случалось наблюдать восходящее перерождение этого пучка на коротком протяжении кверху от места разрушения».
Ввиду всего вышеуказанного нельзя не признать совершенно ошибочным утверждение А. Брюса2, будто бы до сих пор не было сделано подразделения пограничного слоя на составные части. Последний автор, между прочим, опубликовал случай амиотро-фического бокового склероза, в котором вместе с перерождением боковых пирамидных пучков и клеток передних рогов обнаруживалось также перерождение волокон основного пучка передне-боковых столбов спинного мозга и одновременно с тем перерождение волокон передней части пограничного слоя; между тем как волокна его задней части, стоящей в связи с клетками задних рогов, представлялись неперерожденными
Благодаря этому автор описывает заднюю часть пограничного слоя в виде особого пучка, широкий конец которого располагается в углу, образованном передним и задним рогом, задний же конец, постепенно истончаясь, протягивается почти до верхушки заднего рога.
1 Упомянутые здесь рнсункн приведены там же, на стр. 99 «Проводящих путей мозга».
2 А. В г и с е, Revue neurol., 1896, № 23.
72
Сравнивая это описание и рисунки автора, я должен признать, что они относятся к той именно области пограничного слоя, которую я описал уже несколько лет назад под названием «внутреннего пучка боковых столбов».
Таким образом, в исследовании А. Брюса, основывающимся на методе перерождения, я нахожу полное подтверждение впервые выделенного мною по методу развития в задней части пограничного слоя особого внутреннего пучка боковых столбов спинного мозга.
ОБ ОСОБОМ ЯДРЕ СЕТЕВИДНОГО ОБРАЗОВАНИЯ НА УРОВНЕ ВЕРХНИХ ОТДЕЛОВ ВАРОЛИЕВА МОСТА
В последнее время, благодаря исследованиям Роллера, моим, Миславского и др., в сетевидном образовании, кроме уже ранее хорошо известных образований (как например, верхние оливы и пр.), удалось выделить несколько новых или мало известных ранее гнезд серого вещества, в числе которых, придерживаясь восходящего топографического расположения, мы назовем: 1) описанное Миславским ядро переднего столба, или дыхательное ядро, заложенное в продолговатом мозгу на уровне корешков п. hypoglossi частью кнутри, частью кнаружи от последних;
2) нижнее центральное ядро Роллера, расположенное в продолговатом мозгу над головным или верхним отделом нижних олив;
3) описанное мною на уровне моста сетчатое ядро покрышки, расположенной над внутренней частью петлевого слоя; 4) описанное мною верхнее центральное ядро, лежащее по обе стороны шва на уровне заднего двухолмия; 5) описанное мною же безымянное ядро, заложенное на уровне переднего двухолмия снутри и стылу от петлевого слоя, представляющегося на этом уровне на поперечных срезах в виде серповидного пучка, и, наконец, 6) расположенное еще выше небольшое конической формы ядро поперечного пучка мозговой ножки.
Кроме этих гнезд серого вещества, в сетевидном образовании может быть выделено еще одно важное гнездо на уровне моста тотчас позади заднего двухолмия, заложенное в глубоких частях покрышки г.
Ядро это мне удалось выделить прежде всего на последовательном ряде окрашенных пикрокармином срезов из мозгового ствола кошки; у которой оно состоит из редко расположенных многоотростчатых больших нервных клеток, различаемых на
1 Рисунок этого ядра и топографическое отношение его к другим соседним ядрам можно найти в первом томе моих «Проводящих путей мозга», СПБ, 1896, стр. 148, рис. 91, под буквами пс se; об этом ядре я упоминаю в том же издании на стр. 143.
74
окрашенных срезах даже невооруженным глазом в виде мелких точек. Таким образом, по своей величине клетки описываемого ядра ничуть не уступают двигательным клеткам передних рогов спинного мозга, а некоторые из его клеточных элементов даже превосходят последние.
В гистологическом отношении описываемое ядро, состоящее из больших многоотросчатых клеток, заложенных среди белого вещества, довольно близко напоминает собой нижнее центральное ядро и подобно последнему располагается в тех частях сете-видного образования, которые являются продолжением основных пучков боковых столбов. В силу этого ядро это может быть названо наружным верхним центральным ядром, или просто верхним центральным ядром, тогда как описанное мною ранее верхнее центральное ядро в отличие от предыдущего может быть названо внутренним верхним центральным ядром или еще лучше срединным ядром.
Вышеуказанное ядро на соответствующем уровне найдено мною, между прочим, и в мозгу собаки, а также и у человека; но у последнего клетки этого ядра не так значительны по своей величине, вследствие чего и самое ядро обозначается далеко не столь резко, как в мозгах кошки и собаки.
Что касается до связей этого ядра, то пока можно сказать лишь, что подобно нижнему центральному ядру оно располагается на пути волокон, являющихся продолжением основного пучка боковых столбов спинного мозга и по всей вероятности стоит в ближайшем соотношении с волокнами последних.
О РАЗВИТИИ КЛЕТОК МОЗГОВОЙ КОРЫ У ЧЕЛОВЕКА х
(Краткое сообщение)
При исследовании зародышевых мозгов человека по методу Гольджи в заведываемой мною лаборатории удалось хорошо окрасить нервные элементы мозговой коры как в период первоначального образования клеток последней, так и в период дальнейшего их роста и развития. На основании этих препаратов я могу сделать следующие выводы относительно развития нервных клеток мозговой коры.
1. Первоначальному образованию клеток мозговой коры на место зародышевых элементов предшествует рост окрашивающихся по Гольджи в черный цвет особых нитей, resp. волокон, выходящих густыми рядами со стороны также окрашенного в черный цвет поверхностного слоя, углубляющихся в зародышевую мозговую ткань и проникающих между зародышевыми тельцами. В наиболее раннем периоде встречаются лишь небольшой длины первичные нити, проникающие в наиболее наружные отделы мозговой коры, позднее же эти нити представляются уже значительно большей длины и достигают более глубоких ее слоев.
2. Самые клетки обязаны своим развитием тому, что вышеуказанные первичные нити при своем развитии, проникая между зародышевыми тельцами, образуют на своем пути изгибы в виде колена или складки вокруг зародышевых телец. Последние при постепенном увеличении колена или изгиба нитей обволакиваются ими по всей своей окружности, образуя, таким образом, закругленный, чаще всего овальный остов будущей нервной клетки, обращенная же кнаружи часть первичной нити становится верхушечным отростком клетки, являющимся, таким образом, по своему развитию наиболее ранним из всех вообще клеточных отростков.
3. Зародышевое тельце, неокрашивающееся по методу Гольджи, служит при выше описанном развитии нервной клетки
1 Доклад, сделанный в научном собрании врачей клиники душевных и нервных болезней в С.-Петербурге 15/IV 1896 г. с демонстрацией препа* ратов.
76
к образованию ее ядра, вещество же нити, обволакивающее зародышевое тельце, служит к образованию клеточной протоплазмы и ее отростков. Благодаря этому во всех клетках наиболее раннего возраста клеточные ядра представляются совершенно неокрашенными, количество же окружающей его протоплазмы по сравнению с размерами ядра оказывается весьма незначительным.
4. В дальнейшем периоде развития от овального остова тела клетки при их росте выступают в виде толстых шипов боковые отпрыски, развивающиеся в протоплазменные отростки, тела же клеток, благодаря этому, становятся угловатыми или близкими по своей внешней форме к пирамидкам. Вместе с тем и узловатости первичных нитей при дальнейшем росте служат началом шипо-образных выступов, которые со временем становятся боковыми побегами верхушечных протоплазменных отростков. Коллатера-ли осевых цилиндров вырастают всегда позднее самих цилиндров на месте четкообразных вздутий последних.
5. Богатое развитие протоплазменных клеточных отростков и их дальнейших разветвлений есть также продукт позднейшего развития клеток, так как более молодые клетки, имея уже сравнительно хорошо развитый верхушечный отросток, почти вовсе лишены как основных, так и боковых протоплазменных отростков или же имеют лишь весьма ограниченное количество основных коротких отростков (согласно Маджини г, Кахалю2 и др.3). Лишь со временем, вместе с ростом тела клетки, развиваются или удлиняются основные отростки и затем ветвится верхушечный отросток, как это мы видим у вполне развитых клеток.
6. Что касается так называемых грушевидных придатков (appendices pyriformes), то они совершенно отсутствуют у более молодых клеток и развиваются на клеточных отростках лишь в позднейшее время, когда клетки уже приобретают вид взрослых. Ранее всего и притом в большом изобилии они развиваются, по-видимому, на верхушечном отростке как более раннем по своему развитию. С развитием протоплазменных отростков число этих придатков все более и более увеличивается4.
7. Различные области мозговой коры развиваются далеко не в одинаковое время. Тогда как в одних областях, например, в чувство-двигательной, мы встречаем уже ясные зачатки клеток, в других областях (например, в subiculeum аммонова рога) мы
1 М a g i π i, Sur la neurologic et les cellules nerveuses cerebrales chez le foetus, Arch. ital. de biol., IX, p. 59.
2 Ramon у Cachal, Sur la structure de 1' ecorce cerebrale de quel-ques mamumiferes, La cellule, 1891, v. VII.
3 К аналогичным результатам пришла в позднейшее время и Стефанов-ская на основании исследований, произведенных над молодыми крысами (М. S t e f a η owsk a, Evolution des cellules nerveusses corticales, Bru-xelles, 18ОД).
4 M. Стефановская, там же, стр. 33 н следующие.
находим лишь первичные нити без всяких следов образования нервных клеток. Равным образом и клетки различных слоев одной и той же области мозговой коры развиваются неодновременно: клетки более глубоких слоев, например, слоя больших пирамид, развиваются ранее клеток более поверхностных слоев.
8. В общем клетки мозговой коры принадлежат по своему развитию к наиболее поздним клеточным элементам центральной нервной системы, так как они представляются еще в зачаточном состоянии в то время, как клетки мозговых узлов представляются уже сравнительно хорошо сформировавшимися, а клетки спинного мозга уже»вполне развитыми.
II
ТРУДЫ ПО ФИЗИОЛОГИИ
ОБ ОТПРАВЛЕНИИ ЗРИТЕЛЬНЫХ БУГРОВ
В физиологии нервной системы нет более темной области, как отправление зрительных бугров. Различные авторы высказали в этом отношении самые разноречивые мнения, что, повиди-мому, давало возможность предполагать о весьма многосторонней роли упомянутых узлов в мозговой деятельности. Между тем до настоящего времени только отношение этих узлов к зрению может считаться более или менее прочно установленным, что, впрочем, очевидно уже из анатомических данных. Все же остальные указания, сделанные теми или другими авторами, относительно значения зрительных бугров в отправлениях нервной системы оспаривались с самых различных сторон и далеко не могли послужить к действительному выяснению отправления этой важной части мозговых полушарий.
Мы считаем здесь необходимым вкратце упомянуть только о наиболее существенных данных, встречающихся в физиологической литературе зрительных бугров.
Начиная с Роландо, многие авторы (из более новых Ренци1, Нотнагель и Ферье2) упоминают о слепоте противуположного глаза, наступающей у животного вслед за повреждением зрительного бугра, даже и в том случае, если разрушению подверглась только передняя его часть. Вместе с тем, некоторые из физиологов думали также установить влияние зрительных бугров и на движение зрачка. Так, Лонже3 при разрушении бугра наблюдал неподвижность зрачка при прямом раздражении светом. Между тем Ренци указывает, что подобное явление наступает только при разрушении передней части узла, тогда как возможно полное разрушение задней его части вызывает продолжительное и сильное расширение противуположного зрачка. Последнее наблюдал также и Ферье в опыте над обезьяной.
Далее, приводили множество данных, говорящих в пользу влияния зрительных бугров на различные роды чувствительности.
1 Renzi, Saggio di fisiologia, Annalt univers. di medicina, m. 186.
2 Ferrier, Die Funktionen des Gehirns; нем. перев. Obersteiner'a, Braunschweig, 1879, стр. 265 и ел.
8 Longet, Anatomie et physiologie du systeme' nerveux, т. 1, стр. 500 и ел. У
6 В. М. Бехтерев 81
Утверждали, например, что, помимо значения бугра для зрения, он влияет также на слух, обоняние, вкуси кожные ощущения.
Представителями взгляда, допускающего чувствительную функцию зрительных бугров, могут считаться Ренци, Фурнье1, Ферье и в особенности Люи2. Мнение этих авторов основывается, главным образом, на том, что при экспериментальных 'исследованиях, в особенности над низшими животными, вслед за разрушением бугра нередко наблюдали притупление или полное прекращение чувствительности на всей противуположной половине тела. Люи в пользу своего взгляда приводит также несколько патологических наблюдений.
В литературе существуют также весьма многосторонние указания о влиянии зрительных бугров на двигательную сферу. Оставляя в стороне взгляды старых авторов (Сосерот, Серре и др.), локализировавших в этих узлах центры для движения передних конечностей, мы находим много указаний на появление вслед за разрушением бугров различных вынужденных движений. Так, Мажанди3 при своих опытах наблюдал круговые движения на сторону поврежденного узла, а Лонже — в противуположной направлении; между тем Шифф4 утверждает, что только повреждение задней части бугра, подобно повреждению мозговой ножки, вызывает кружение животного на здоровую сторону (согласно с мнением Лонже); повреждение же передней трети узла сопровождается кружением животного в противупо-ложную сторону (согласно синением Мажанди). Впрочем указания Ренци более согласуются с мнением Лонже. Кроме того, Ренци в опытах над птицами с разрушением мозговых полушарий вместе с зрительными буграми наблюдал полную неспособность животных стоять, ходить и летать, несмотря на то, что они могли еще производить отдельные движения и махали крыльями при бросании их на воздух.
Наконец, не очень давно были опубликованы опыты Нотнаге-ля (над кроликами), в которых разрушение бугров производилось впрыскиванием хромовой кислоты. Существенные результаты этих опытов состояли в том, что оперированные животные не представляли никаких явлений двигательного или чувствительного паралича, но. отличались от здоровых тем, что при пассивном вытягивании или передвижении их передних конечностей-оки долгое время сохраняли новые положения своих членов. Принимая, что исправление положения членов у нормальных животных выполняется отраженным путем под контролем мышечного чувства, ав-
1 F о u r η i ё, Recherches experim. sur le fonctionnement du cerveau, Paris, 1873.
2 Luys, Recherches sur le systeme nerveux, стр. 538.
3 Magendie, Legons sur les fonctions etc., Paris, т. I, 1839.
4 Schiff, Lehrbuch der Physiologie des Menschen, т. I, 1858—1859.
82
тор приходит таким образом к заключению об отраженном отправлении зрительных бугров.
Нетрудно видеть, что, на основании столь разноречивых данных, нет никакой возможности дать себе правильный отчет об явлениях, следующих за разрушением зрительных бугров. Таким образом, необходимость новых исследований, с соблюдением самой строгой осмотрительности при вскрытии оперированных животных, становится неизбежно, если в занимающем нас вопросе желают достигнуть хоть сколько-нибудь удовлетворительных результатов.
С этой целью я предпринял ряд опытов над различными животными: лягушками, голубями, курами, кроликами и собаками. Полученные мною результаты, как мне кажется, в достаточной мере уясняют значение зрительных бугров в отправлениях нервной системы вообще.
Во многих опытах я пользовался способами как раздражения, так и разрушения. Опыты с разрушением зрительных бугров делались по 2 принципам: в одних разрушение производилось без предварительного разрушения мозговых полушарий. В других же — над менее высоко стоящими животными (лягушками и птицами) — предварительно, тем или иным способом, удалялись полушария головного мозга, и только затем уже подвергались разрушению зрительные бугры, чтобы определить, какие отправления из числа тех, которые еще оставались у животного после удаления мозговых полушарий, исчезают вместе с разрушением зрительных бугров.
Так как при своих опытах, между прочим, я стремился проверить и существовавшие до сих пор в литературе мнения относительно отправления и значения зрительных бугров, то прежде, чем изложить существенные результаты своих опытов, я скажу сначала, к каким выводам привели эти опыты в указанном отношении.
Что касается до отношения бугров к зрению, то, ввиду несомненно доказанного количественного различия в перекресте зрительных волокон в chiasma у различных видов животных, мне казалось наиболее важным и интересным произвести опыты над вышестоящими животными, над которыми, сколько мне известно, до сих пор еще не было производимо систематических исследований подобного рода. Один опыт над обезьяной, приводимый Ферье, конечно, не может считаться в этом отношении достаточным,— тем более, что в этом опыте одновременно с зрительным бугром было произведено и разрушение gyri angula-ns (коркового зрительного центра).
В опытах над собаками я прежде всего убедился, что разрушение, как передней и в особенности средней части узла, так и его заднего отдела, вызывает расстройства со стороны зрения. Но при разрушении передних отделов узла расстройства со стороны зрения были в высшей степени кратковременны и нередко
6* 83
уже по истечении 1—2 дней исчезали настолько, что не могли быть обнаружены, даже при внимательном исследовании. При разрушении же заднего отдела узла нарушение зрительной способности животного представлялось более или менее стойким и могло быть обнаружено даже по прошествии целых месяцев после операции. В таких случаях у животных по большей части наблюдалось половинное ограничение поля зрения на сторонах про-тивуположных в отношении разрушенного бугра (исчезновение отправления одноименных половин сетчаток на соответствующей стороне). Иногда, впрочем, при меньших повреждениях слепота ограничивалась половиной сетчатки только одного глаза. Таким образом, результаты моих опытов в этом отношении находятся в полном согласии с недавно полученными мною фактами, несомненно говорящими в пользу существования неполного перекреста зрительных волокон в chiasma собаки 1.
Что касается до указаний авторов о влиянии бугров на состояние зрачков и происхождение различных вынужденных движений и расстройств в равновесии тела, то, хотя в некоторых моих опытах, действительно, и наблюдались явления, сходные с теми, о которых упоминают авторы, но я убедился, что они не зависели от разрушения самой массы бугра, а находились в связи с одновременным повреждением прилежащего отдела центрального серого вещества, окружающего наиболее глубокую часть 3-го желудочка2.
Относительно перемен в чувствительной сфере мои опыты с разрушением бугров у собак привели к совершенно отрицательным результатам. То же я должен сказать и о наблюдении Нотнагеля касательно сохранения животными положения своих членов при пассивном их передвижении. Наконец, я не наблюдал у оперированных мною животных и никаких явлений двигательного паралича конечностей.
На основании этих отрицательных данных я прихожу к заключению, что все подобного рода явления, упоминаемые в опытах авторов, не составляют последствий разрушения самого бугра, а, по всей вероятности, находятся в зависимости от совместного повреждения волокон, проходящих в соседней части внутренней сумки.
Таким образом, из всех имеющихся в литературе указаний относительно значения зрительных бугров наши опыты подтверждают только непосредственное отношение их к зрению. Но естественно, что нельзя удовольствоваться одним влиянием зрительных бугров на зрение, так как опыты нам показывают,
1 См. мою работу «О перекресте зрительных" волокон в chiasma ηη. opticorum» в еженедельной клинической газете и в Neurologisches Zentralblatt за 1883 г.
2 См. мою работу: «Die Bedeutungder Trichterregion des 3 Ventrikels f. die Erhaltung des Korpergleichgewichtes» в St.-Petersb. Med. Wochen-schrift, N 12, 1882. О влиянии центрального серого вещества на состояние зрачка было сделано мною сообщение в Обществе психиатров в 1882 г.
84
что только' повреждение незначительной их части — именно задней (может быть, даже исключительно одних corpora geniculate) — сопровождается стойким нарушением зрительной способности. В настоящее время вообще признано, что название зрительного бугра меньше соответствует собственно бугру, нежели corpus quadrigeminum или bigeminum.
Нижеследующие факты, которые мне удалось получить при опытах над различными животными, могут пополнить существовавший до сих пор пробел относительно отправления зрительных бугров.
До сих пор утверждали, будто зрительный бугор вовсе невозбудим электрическими токами; но я нередко замечал, что при механическом повреждении этого узла, как у высших животных, так и у низших, получались те или другие, свойственные этим животным, крики. Голуби при указанной операции нередко произносят протяжный звук «гуль, гуль», похожий на воркование; курица издает своеобразный крик; собака начинает визжать, лаять или ворчать. Такого же рода результаты, но только с большим постоянством, получались и при раздражении зрительных бугров фарадизационным током *. У птиц своеобразные крики получались даже и при раздражении бугров после предварительного удаления мозговых полушарий; кроме того, в опытах над собаками я мог убедиться, что вслед за перерезкой на уровне мозговых ножек, тотчас позади зрительных бугров, раздражение последних уже не в состоянии более вызывать своеобразных криков животного. Все эти данные заставили меня подозревать, что зрительные бугры служат центрами таких движений, при помощи которых выражается эмоциональная сторона душевной жизни. Чтобы проверить это предположение, я и произвел мои позднейшие опыты.
Известно, что еще Лонже и затем Вульпиан2 обратили внимание на тот факт, что кролики, лишенные мозговых полушарий, способны обнаруживать крик под влиянием сильных болевых раздражений. Так как эти авторы вместе с тем убедились, что перерезка варолиева моста прекращает жалобный крик животного, вызываемый болевыми раздражениями, то они и локализировали в варолиевом мосту центр, при помощи которого животное становится способным выражать ощущения боли.
Вслед за тем Гольц, производя опыты над лягушками, показал, что вслед за удалением мозговых полушарий животное квакает каждый раз после механического раздражения кожи на боковых частях спины; рассечение же мозговой ткани в области четверохолмия (собственно-двухолмия) прекращало этот звук
1 С указанной целью я пользовался тонкими, проволочными электродами, уединенными на всем своем протяжении, за исключением самых окончаний, при помощи особенного дурно проводящего состава; электроды эти погружались в мозговую ткань до массы узла.
Vulpian, Lecons sur la physiologie gener. et сотр., Paris, 1865.
85
навсегда. На этом основании автор локализировал отраженный центр кваканья лягушки в четверохолмии Ч
Наконец, в недавнее время Ферье, основываясь на своих опытах, высказал гипотезу, по которой задние отделы corporum quadrigeminum (а у нижестоящих животных corpus bigeminum) служат центром для отраженного выражения ощущений и чувствований. Фактические основания этого взгляда, однако, не могут считаться достаточными. Автор основывает свое заключение, главным образом, на опытах с раздражением заднего отдела четверохолмия, причем у животного, кроме различных судорожных явлений (расширение противуположного зрачка, отклонение головы и глаз вверх и в противупрложную сторону, оттягивание уха, сжатие челюстей, стягивание углов рта и, наконец, общий opisthotonus), замечались еще и различные крики, от короткого слабого стона — при слабом действии электродов — до самых различных, свойственных оперированным животным, криков — при продолжительном и сильном раздражении. Напротив, опыты с разрушением этой области (которые, кстати сказать, должны бы иметь гораздо больше цены, нежели опыты с раздражением), повидимому, не привели к удовлетворительным результатам, так как автор выражается по этому поводу следующим образом: «если же четверохолмие удалялось совершенно при помощи поперечного разреза, то спустя 2—3 часа после операции кожные раздражения, достаточные, чтобы вызвать отраженные движения членов, не могли уже возбудить никакого крика, что все-таки удавалось позднее при сильных кожных р а з д р а ж е н и-я х» 2.
Если принять во внимание только одни факты, сообщаемые цитированными авторами, то, мне кажется, что их исследования нисколько не противоречат нашему предположению об отправлении бугров, ибо несомненно, что отраженные волокна, подходящие к каждому бугру, должны проходить через нижележащие области (продолговатый мозг и варолиев мост) и что раздражение этих волокон в указанных областях может вызывать отраженные крики, тогда как глубокая перерезка, производимая в той или иной части моста, должна привести к совершенной неспособности животного выражать свои ощущения криками.
Таким образом, мне оставалось только найти фактическое подтверждение сделанному мною предположению об отправлении зрительных бугров.
Я много раз производил замечательный опыт Гольца над кваканьем лягушки и убедился, что разрез мозга, производимый Гольцем у этих животных, должен проходить впереди зрительных бугров, оставляя последние совершенно нетронутыми.
1Goltz, Beitr3ge~ z. Lehre v. d. Funktionen d. Nerven zentren des Frosches, Berlin, 1869.
2 Ferrier, Die Funktionen des Gehirns, стр. 86.
86
Чтобы убедиться, что двухолмие в этом опыте не играет никакой роли в произведении отраженного кваканья, как это утверждает Гольц, я вскрываю вслед за тем черепную покрышку лягушки и, осторожно захватив тонким пинцетом поочередно каждое из возвышений обнаженного двухолмия, постепенно раздавливаю их между ножками пинцета до совершенного уничтожения. Спустя немного времени после подобной операции кваканье животного снова удается вызвать, раздражая кожу спины. Если же вслед затем в области разрушенного двухолмия провести глубокий разрез, который должен таким образом проходить позади зрительных бугров, то кваканье прекращается навсегда, и его не удается уже вызвать никакими способами.
Подобные же опыты я мог произвести и на многих других животных. В особенности пригодны для этого различные птицы (воркующие голуби, куры и пр.), выражающие свои ощущения самыми разнообразными криками.
Разрушая голубю мозговые полушария с сохранением зрительных бугров, я убедился, что вслед затем у такого голубя удается вызвать крик «гуль, гуль» каждый раз, когда его поднимают на воздух за одно крыло и когда он начинает сильно биться. Подобный же крик у оперированного голубя удается вызвать и при помощи быстрого щипка за кожу верхней части шеи. Другие кожные раздражения (за исключением крайне болезненных) не в состоянии вызвать этого крика. Затем я разрушал у такого голубя corpora bigemina и мог получать тот же своеобразный крик при вышеуказанных условиях. Точно так же разрушение обоих corpora bigemina у здоровых голубей никогда не нарушает способности их издавать крики. Напротив, воркующие голуби, которым я производил разрушение бугров, как при сохраненных мозговых полушариях, так и после предварительного удаления их навсегда утрачивали способность издавать звуки, которые не удавалось уже вызывать даже и сильными болевыми раздражениями.
Подобные же опыты, с совершенно тождественным результатом, я производил на курах, у которых вслед за удалением мозговых полушарий с сохранением зрительных бугров своеобразные крики могли быть вызываемы с неменьшим постоянством, как у безмозглой лягушки кваканье, механическим раздражением боковых частей спины, щипком кожи в верхней части шеи или даже быстрым захватыванием перьев в той же области. Разрушение corp. bigemina в этих опытах также не нарушало машинообразных криков животного, которые прекращались только вслед за разрушением зрительных бугров.
Из высших животных, бывших в моем распоряжении, собаки оказались наиболее пригодными для опытов с указанной целью, так как, кроме разнообразных криков, они обнаруживают свои ощущения еще различными выразительными движениями и способны даже к мимической игре мышц лица, которую у них легко
87
вызвать щипком за кожу щеки или за крыло носа или сильными вкусовыми и обонятельными раздражениями.
Прежде всего я убедился, что и у собак после весьма значительного разрушения поверхности мозговых полушарий легко удается вызвать отраженные крики (визг, лай, ворчание) при помощи сильных болевых раздражений. Раздражения кожи лица или вкусовых нервов вызывали мимическую игру мышц на стороне раздражения (сморщивание носа, оскаливание зубов). Особыми опытами я мог также убедиться, что изолированное разрушение четверохолмия нисколько не нарушало способности животного выражать свои ощущения.
При разрушении одного бугра или при перерезке тотчас позади его животное в состоянии еще произносить звуки под влиянием болезненных ощущений, хотя, повидимому, и значительно слабее, нежели здоровое животное, но мимические движения лица на стороне, противуположной разрушенному бугру, прекращаются совершенно. Их не удается вызвать даже и сильным щипанием кожи щеки или крыла носа, за которым наступают только слабые движения с характером простых рефлексов. При этом никакой асимметрии в покойном положении лица не замечается, и собака в состоянии произвольно сокращать мышцы обеих сторон, что можно заметить, например, при облизывании животного. Такие животные, даже и после того, как они оправятся от операции, становятся вялыми и не выражают своих ощущений различными движениями и звуками с той живостью, как это делают здоровые собаки.
С разрушением обоих зрительных бугров или при глубокой перерезке мозговых ножек у собак совершенно прекращаются те разнообразные, отраженные крики, которые всегда обнаруживают нормальные животные при болевых раздражениях кожи. Только крайне болезненные раздражения иногда вызывают у них еще короткий звук, похожий на звуки, которые можно вызвать у собак сильным и быстрым надавливанием на грудную клетку.
Опыты над кроликами с разрушением обоих зрительных бугров привели к подобным же результатам, как и опыты на собаках.
Итак, из всех вышеприведенных данных очевидно, что животные, лишенные мозговых полушарий, если у них сохранены в целости зрительные бугры, могут еще под влиянием внешних раздражений издавать те разнообразные крики, которыми нормальные животные пользуются для выражения своих ощущений. Эти крики удается вызвать у животных даже и в том случае, если вслед за удалением мозговых полушарий подверглись разрушению и возвышения corp. quadrigeminum. Только после разрушения зрительных бугров способность животного издавать отраженные крики прекращается навсегда. Животные же, у которых полушария головного мозга оставлены в целости и
88
разрушены только одни зрительные бугры, сохраняют полную свободу своих движений и, очевидно, еще в состоянии правильно воспринимать ощущения, но они теряют возможность обнаруживать эти ощущения при помощи соответствующих движений и криков.
Таким образом из наших опытов следует, что в зрительных буграх сосредоточены приводы для непроизвольной иннервации тех мышечных групп, которые участвуют в выражении разнообразных ощущений, свойственном данному виду животных. Эти узлы, следовательно, по преимуществу служат для так называемых выразительных криков и движений; четверохолмие же, в котором новейшие авторы локализировали центры для выражения ощущений и чувствований, не играет в этом отношении никакой роли.
Из наших опытов очевидно также, что если у животного удалены мозговые полушария, то возбуждение зрительных бугров к деятельности может происходить только отраженным путем вследствие раздражений, действующих на чувствующую поверхность тела. В здоровом же состоянии животного это возбуждение, как легко понять, может происходить также и под влиянием психических импульсов, которые, однако же, могут не иметь, ничего общего с произвольными побуждениями.
Работу свою я не считаю вполне законченной и более подробное опубликование результатов всех сделанных мною опытов^ равно как и всестороннее обсуждение их, я оставляю до бли-жейшего будущего.
ОБ ИННЕРВАЦИИ И МОЗГОВЫХ ЦЕНТРАХ СЛЕЗООТДЕЛЕНИЯ
Проф. В.Бехтерев и д-рН. Миславский
Первые указания на отделение слез под влиянием возбуждения нервов мы находим у Чермака х. Раздражая общий ствол п. trigemini на отрезанной и распиленной пополам голове кролика, Чермак наблюдал появление слез в соответствующем глазу. Вместе с этим он заметил увеличение отделения конъюнктиваль-ных желез, а равно и гардеровой железы. Этим, в сущности, и ограничивались наши сведения об иннервации слезных желез до работы Герценштейна 2, появившейся в 1868 году. Последний автор, оперируя над собаками, убедился, что раздражение n.lacri-malis и subcutanei malae вызывает обильное отделение слез, которое может быть получено даже после перевязки сонной артерии соответствующей стороны. Точно так же раздражение чувствующих 1-й и 2-й ветвей п. trigemini в опытах Герценштейна вызывало отделение слез на соответствующей раздражению стороне. С другой стороны, известно, что слезоотделение появляется и при сильном световом раздражении сетчатки глаза. Что касается шейного ствола симпатического нерва, то перерезка его остается без влияния на отделение слез, раздражение же этого нерва в опытах Герценштейна дало столь изменчивые результаты, что автор не решился высказаться на основании их ни за, ни против влияния этого нерва на отделение слез. Следует, далее, отметить в опытах Герценштейна тот факт, что перерезка п. 1а-crimalis не вызывала остановки или уменьшения обыкновенного более или менее постоянного отделения слез. Впрочем, продолжающееся после этой перерезки отделение слез Герценштейн считает за паралитическое. С другой стороны, отравление животного кураре даже после перерезки п. lacrimalis вызывало в его опытах некоторое увеличение слезоотделения, которое представлялось более умеренным на стороне произведенной перерезки, нежели в противоположном глазу.
1 Czermak, Molleschott's Untersuchungen, 1860, Bd. VII, S. 379. s Herzenstein, Beitrag zur Physiologie und Therapie der Tr3-nenorgane, Berlin, 1868.
90
Данные Герценштейна были проверены и расширены наблюдениями Демченко х. Опыты этого автора были произведены на наркотизированных морфием собаках, кошках и кроликах. Подтвердив Герценштейна по вопросу о влиянии раздражения п. lacrimalis на отделение слез, Демченко отрицает слезоотделительное значение п. subcutanei malae. В его опытах лишь иногда вслед за раздражением этого нерва глаз казался несколько влажным, но и то в такой степени, что трудно было сделать какие-либо положительные заключения. С другой стороны, обнаруживающееся вслед за перерезкой п. lacrymalis слезотечение Демченко объясняет параличом век вследствие неизбежного при операции повреждения нервов, заведующих движением последних. Что касается до влияния п. sympathici, то Демченко с положительностью утверждает, что раздражение этого нерва вызывает ясное отделение слез, безразлично, был ли при этом п. lacrimalis предварительно перерезан или же оставался невредимым. Наконец, рефлекторное слезоотделение Демченко получал при раздражении ηη. frontalis infraorbitalis lingualis, glossopharyngeus и vagus (?) 2. У людей при раздражении носовой полости появлялось слезоотделение сначала на соответствующей стороне, а несколько позднее и на противоположной. Из спинномозговых нервов можно было получить отделение слез с п. auricularis magnus, хотя и слабее, чем с вышеупомянутых го-ловно-мозговых нервов; между тем с п. dorsalis pedis рефлекс уже не получался. Наблюдения на людях точно так же будто бы показали, что рефлекторное отделение слез возможно при операциях на голове и лице; при операциях же на туловище и конечностях (даже без наркоза) рефлекторного отделения слез не наблюдается. Следует еще заметить, что рефлексы получаются только при целости п. lacrimalis, с перерезкой же его они пропадают. Напротив того, перерезка п. sympathici остается без влияния на рефлекторное слезоотделение.
Вольферс3, работа которого появилась почти одновременно с работой Демченко, также нашел, что п. lacrimalis и п. sympathies при раздражении вызывает отделение слез, причем sympathies обнаруживает действие после перерезки п. lacrimalis, следовательно, волокна его направляются отдельным путем к железе. По Вольферсу, также и п. subcutaneus malae действует на железу, вызывая отделение слез, которое обнаруживается сильнее у овец; у собак же эффект раздражения этого нерва проявляется слабее. Что касается рефлексов, то Вольфер-сом были изучены только рефлексы с ветвей п. trigemini. При этом он убедился, что целость ηη. lacrimalis и subcutanei ma-
1 Демченко, Z. Innervation der Tranendrflse, Pfluger's Archiv, 1872, Bd. VI, S. 191.
1 Вопросительный знак в подлиннике.
* Wolfe rs, Experimentelle Untersuchungen fiber die Innerva-tionswege der Tranendrflse. Dissert., Dorpat, 1871.
91
lae необходима для получения рефлексов. Между тем опыты с влиянием самих перерезок ηη. lacrimalis] и subcutanei malae Вольферс считает мало доказательными ввиду больших повреждений, связанных с этими операциями.
Что касается до влияния центральных частей нервной системы на слезоотделение, то в литературе мы нашли только указание Экхарда, по которому центр для отделения слез помещается в верхнем отделе шейной части спинного мозга, начиная с 5-го шейного позвонка до начала п. trigemini.
Таким образом, центральная иннервация слезных желез остается до сих пор наименее всего изученной, а влияние высших мозговых центров на деятельность этих желез и совершенно неизвестным. Между тем отделение слез связано теснейшим образом с психической деятельностью и служит, как известно, наиболее характеристичным выразителем разнообразных душевных волнений. Ввиду этого мы и приступили к изучению локализации высших мозговых центров слезоотделения и тех путей, посредством которых передается возбуждение этих центров на слезоотделительные нервы.
Опыты наши производились на собаках, умеренно кураризи-рованных. Мы должны были прибегнуть к кураризации животных ввиду того, что судороги, развивающиеся при раздражении коры и подкорковых центров, неизбежным образом нарушали бы чистоту наблюдения. К тому же из своих опытов мы убедились, что усиление отделения слез при кураризации наступает далеко не всегда в такой степени, чтобы могло затруднять наблюдения, а нередко его даже и вовсе не обнаруживается.
Что касается до способа наблюдения, то мы в своих опытах следили за отделением слез по преимуществу непосредственно глазом, слегка оттянувши нижнее веко; при этом мы считали за несомненный эффект только те результаты раздражения, при которых наблюдалось обильное отделение слез. Небольшие же изменения во влажности глаз мы не считали доказательными в каком-либо отношении уже потому, что они могут зависеть и не от выделения слезных желез. По крайней мере из опытов и наблюдений Бернарда, Гекстора, Фрерикса и Демченко видно, что и после вылущения этих желез глаз все же представляется увлажненным, да и некоторое увеличение влажности глаза, как показал Чермак, может зависеть от увеличенного отделения конъюнктива л ьных желез.
Трепанация и удаление верхней части полушария для исследования центров основания мозга в наших опытах выполнялись обычным порядком, раздражение же производилось ясно ощутимым на языке током, взятым от санного аппарата Дюбуа-Реймона с одним элементом Грене.
Прежде всего путем особых опытов мы убедились в несомненном влиянии шейного ствола п. sympathici на слезную железу. В наших опытах при раздражении периферического конца
92
η. sympathici вместе с расширением зрачка, выпячиванием глаза и уходом третьего века кнутри обнаруживалось вполне ясное слезоотделение в соответствующем глазу. Нечего и говорить, что раздражение п. trigemini всегда вызывало обильное отделение слез на стороне раздражения.
Что касается до рефлекторного отделения слез, то, подтверждая в общем указания, сделанные нашими предшественниками, мы должны указать на то обстоятельство, что в наших опытах раздражение центрального конца п. ischiadici всегда вызывало слезоотделение, сопровождаемое заметным выпячиванием глаз, расширением зрачков и уходом третьих век внутрь. При этом слезоотделение хотя и обнаруживалось в обоих глазах, но в противоположном оно всегда наступало несколько ранее и было сильнее, нежели в соответствующем глазу.
Мы не можем далее согласиться с Демченко и Вольферсом, что для получения рефлекторного действия безусловно необходима целость п. trygemini (ηη. lacrimalis и subcutanei malae). Хотя и не во всех опытах, но мы получали не один раз ясное рефлекторное отделение слез в обоих глазах, несмотря на предварительную одностороннюю перерезку п. trigemini. При этом слезоотделение представлялось даже обильнее на стороне перерезанного п. trigemini при условии, если раздражение чувствительного нерва (п. ischiadici) производилось на той же стороне.
Перейдем теперь к изложению результатов нашего исследования в отношении мозговых центров.
Из опытов выяснилось, что в отношении отделения слез наиболее деятельными областями мозговой коры являются внутренние части переднего и заднего отдела сигмовидной извилины. Несколько слабее действуют в этом отношении более кнаружи расположенные отделы той же извилины. С других же областей мозговой коры мы не могли в своих опытах получить вполне ясного отделения слез.
При раздражении вышеупомянутых деятельных областей мозговой коры слезоотделение наступает весьма быстро вслед за началом раздражения, и притом оно часто обнаруживается настолько сильно, что слезы выливаются наружу. Обыкновенно слезоотделение наблюдается в обоих глазах, но почти всегда оно сильнее выражено на стороне, противоположной раздражению. Вместе с прекращением раздражения коры слезоотделение вскоре приостанавливается совершенно, и если затем глаз осушить пропускной бумажкой, то в дальнейшем периоде он представляет уже нормальную степень влажности.
Следует здесь заметить, что вышеописанный эффект в отношении отделения слез при раздражении мозговой коры обыкновенно сопровождается резким двусторонним расширением зрачков, выпячиванием обоих глаз и уходом третьих век кнутри. Впрочем, в противоположном глазу только что упомянутые яв-
93
ления наступают несколько раньше и, повидимому, обнаруживаются с большей резкостью, нежели в соответствующем глазу *.
Исследование мозговых узлов показало, что центры, вызывающие отделение слез, заложены в зрительных буграх. Раздражение одного из этих образований на весьма ограниченном пространстве в глубине внутренней его части, на уровне серой спайки, вызывает слезоотделение в обоих глазах, хотя и здесь эффект всегда обнаруживается с большей силой на стороне, про-тивополжной раздражаемому зрительному бугру.
Как и при раздражении мозговой коры, слезоотделение, вызванное раздражением зрительного бугра, сопровождается расширением зрачков, уходом третьих век кнутри и выпячиванием глаз 2.
Таким образом, из приведенных опытов очевидно, что при раздражении вышеуказанных областей мозговой коры и зрительных бугров представляются заинтересованными центральные продолжения шейного симпатического нерва.
Для того чтобы выяснить, в какой степени эти центральные симпатические волокна оказывают влияние на слезоотделение при раздражении мозговой коры и зрительных бугров, мы в особых опытах производили предварительную одностороннюю перерезку шейного ствола п. sympathies
Как и можно было ожидать уже a priori, в этом случае при раздражении коры и бугров отделение слез обнаруживалось на обоих глазах, хотя расширения зрачка, выпячивания глаза и ухода третьего века кнутри, само собою разумеется, более уже не наблюдалось на стороне перерезки.
В других опытах, поставленных с целью уяснения интересующего нас вопроса, мы производили перерезку п. trigemini внутри черепа перед gangl. Gasseri, оставляя оба п. sympathici в целости. И в этом случае раздражение коры противоположного мозгового полушария вызывало слезоотделение на обоих глазах, хотя на стороне перерезки оно выражалось гораздо слабее. Тот же эффект в этом случае вызывался и раздражением зрительных бугров.
В одном опыте после перерезки п. trigemini мы первоначально не получили отделения слез на стороне перерезки при раздражении коры и зрительного бугра противоположной стороны, но
1 Считаем здесь необходимым заметить, что в одном случае у молодого щенка (приблизительно 2—3-месячного возраста) мы не получили с коры ни отделения слез, ни расширения зрачков, ни выпячивания глаз и ухода третьих век. Интересно, что и симпатический нерв в этом случае оказался совершенно бездеятельным как в отношении слезоотделения, так и в отношении расширения зрачка, выпячивания глаза и ухода третьего века, между тем раздражение п. trigemini внутри черепа вызывало ясное слезоотделение.
2 В вышеупомянутом опыте над молодым щенком мы не получили никакого эффекта не только при раздражении мозговой коры, но и при раздражении зрительных бугров.
94
при этом не наблюдалось и изменений в зрачке, выпячивания глаз и ухода третьего века кнутри; увеличив же несколько силу тока и повторив раздражение зрительного бугра, мы и в этом случае на стороне перерезки получили отделение слез вместе с расширением зрачка, выпячиванием глаза и уходом третьего века кнутри.
Ввиду вышеизложенных данных мы приходим к выводу, что возбуждение мозговой коры и зрительных бугров влияет на слезоотделение как при посредстве п. trigemini, так, по крайней мере отчасти, и при посредстве шейного ствола симпатического нерва.
Из наших опытов выяснилось также, что рефлексы с п. ischiadic! на слезную железу пропадают совершенно после перерезки позади зрительных бугров. Вместе с тем прекращается рефлекторное расширение зрачков и выпячивание глаз, а оба третьих века представляются парализованными.
На основании этих данных следует допустить, что главный рефлекторный центр слезоотделения помещается в зрительных буграх и что там же заложены центральные приводы шейного ствола симпатического нерва, откуда продолжения их поднимаются к коре полушарий.
О ВЛИЯНИИ КОРЫ БОЛЬШОГО МОЗГА НА АКТ ГЛОТАНИЯ И НА ДЫХАНИЕ
Проф. В. М. Бехтерев и д-р П. А. Останков
Сколько нам известно, в литературе имеются лишь крайне скудные указания относительно влияния коры большого мозга на акт глотания. Хотя глотание возможно и в бессознательном состоянии, а равно и после разрушения большого мозга, мозжечка и варолиева моста \ но вместе с этим, конечно, не устраняется и возможность влияния коры большого мозга на акт глотания, которое представляется более чем вероятным не только ввиду участия нашей воли в глотательном акте, но и ввиду непосредственного вызывания глотательных движений вкусовыми представлениями.
Рефлекторный глотательный центр продолговатого мозга, по Ландуа, помещается на дне четвертого желудочка. Более точное топографическое описание этого центра мы имеем в работе Макса Марквальда «О распространении возбуждения и задержки с центра глотания на центр дыхания» 2. «С целью более точного определения положения центра глотания,— пишет Марквальд,— я делал ряд поперечных перерезок на различных уровнях продолговатого мозга, разрушая вместе с тем отдельные части мозга. Отделение продолговатого мозга на высоте tuber-cula acustica прерывает связь между двигательным путем trigeminus и глотательным центром, причем уничтожается первый •отдел глотательного акта — сокращение m. mylohyoidei, в то (время как самое глотание еще возможно; если же с перерезкой итти далее, приближаясь к вершинам alae cinerae, то мы достигнем уровня, по отделении которого глотание не вызывается ни раздражением мягкого неба, ни раздражением п. laryngei superiors; гортань, глотка и пищевод лишаются движения, и глотательный акт уничтожается, в то время как дыхание остается неизмененным».
Только что указанный рефлекторный глотательный центр возбуждается, по Ландуа, при посредстве чувствующих ветвей
1 Landois L., Lehrbuch der Physiologie des Menschen, 1889.
2 Marckwald, Ueber die Ausbreitung dor Erregung und Hemmung vom Schluckzentrum auf das Atemzentrum. Separat. Abdr. aus. der Zschr. f. Biologie, Bd. XXV, Hft. VII, S. 46 — 47.
S6
рта, неба и зева (2 и 3 ветви п. trigemini, glossopharyngeus и чувствующие волокна vagi); центробежный же его путь лежит в двигательных ветвях глоточного сплетения.
В настоящем кратком сообщении мы коснемся лишь влияния коры большого мозга на акт глотания. В этом отношении заслуживают внимания указания, содержащиеся в работе Крау-зе1, который при раздражении слабым током gyr. praefronta-lis теменной доли в «своем вниз направленном побеге» получал повторные глотательные движения, сокращения передней мускулатуры шеи с поднятием гортани, мягкого неба, сжимателей глотки, задней части спинки языка и arcus palato-glossi. В вышеназванной части мозговой коры, по автору, содержится область чувствующей сферы как для передней мускулатуры шеи, так и для движений гортани и глотки. При этом нелишне заметить, что при удалении этой части коры, как и следовало ожидать, не обнаруживалось никаких расстройств в глотательных движениях.
При своих опытах мы пользовались методом, близким к тому, который применялся при исследовании различных вопросов, касающихся акта глотания, Г. Кронекером и С. Мельцером2. Собаке, наркотизированной смесью хлороформа с эфиром, вводился в полость глотки тонкий резиновый баллон, навязанный на конец обыкновенного желудочного зонда. Животное предварительно было трахеотомировано на случай искусственного дыхания. Баллон вводился в пищевод и затем, путем обратного движения (для лучшего расправления баллона), помещался при верхнем его конце. Другой конец желудочного зонда после предварительного наполнения его вместе с баллоном водой соединялся с водяным манометром, второе колено которого связывалось с воздушным мареевским прибором. Таким образом, перо последнего и отмечало толчки и давление, испытываемое баллоном при глотательных движениях животного в течение опытов.
Что касается остальной обстановки последних, то она состояла в том, что после введения баллона в верхний конец пищевода мы трепанировали животному переднюю часть черепной покрышки, после чего твердая мозговая оболочка разрывалась крестообразно и отворачивалась на края костной раны. Для раздражения мозговой коры мы пользовались двумя тонкими изолированными электродами санного аппарата Дюбуа-Рей-мона с элементом Грене.
1 Krause, Du Bois-Reymond's Acrh. f. Anat. u. Physiol., 1884, S. 203.
"Kronecker H. und Meltzer S., Der Schluckmechanis-mus, seine Erregung und seine Hemmung, Arch, fur Anatom. und Phy-siologie, 1883; Ueber den Schluckmechanismus und dessen nervose Hem-mungen, Monatbericht der Konigl. Akadem. der Wissenschaften zu Berlin, 1881.
В. М. Бехтерев
97
Поставленные нами опыты дали следующие результаты.
Раздражая различные участки корковой поверхности мозга у собаки, первоначально мы не получали никаких результатов, т. е. вовсе не замечали движений рычага мареевского прибора; но уже в первый же день исследования (вначале июля 1893 года) мы имели возможность отметить область коры, вблизи переднего края второй борозды (считая снутри), при раздражении которой получалась вполне ясно выраженная кривая глотательного акта (рис. 1). Во всех других исследованных нами случаях область эта всегда занимала тождественное положение на мозговой поверхности как того, так и другого полушария. При этом считаем нужным заметить, что при раздражении вышеуказанной области коры не только получалась кривая глотательного движения, но

Рис. 1.
и ad oculos можно [было убедиться, что животное производило глотание, тем более что последнее обыкновенно сопровождалось характеристичным звуком. При перемещении электродов на соседние участки мозговой поверхности эффект быстро исчезал.
Что касается более точной локализации открытого нами глотательного центра, то наши опыты показывают, что он расположен на передне м отделе второй извилины, кнаружи от сигмовидной извилины, у самого переднего конца второй| борозды (см. а на рис. 2).
Если борозду sulci cruciati мысленно продолжить далее кнаружи, то эта линия проходит как раз через ту область, которая содержит глотательный центр. Последний находится, таким образом, в ближайшем соседстве с открытыми Ферье центрами для движений углов рта, для движения челюстей и языка и с так называемым «ротовым центром» (Mundzentrum), служащим для открывания рта и передвижения языка (по Ферье). Мы убедились, кроме того, что в непосредственном соседстве с открытым нами глотательным центром помещается и выдыхательный центр (см. ниже).
Раздражением основных частей мозга также удается вызвать глотательный акт. Так, в наших опытах при погружении электродов в область зрительного бугра после предварительного удаления большей части полушария вызывались с постоянством глотательные движения. Кроме того, в одном случае при раздражении основных частей мозга тем же способом нам удалось вы-
98
звать весьма бурное рвотное движение. Нам остается еще упомянуть, что глотательный акт в наших опытах вызывался и реф-лекторно электрическим раздражением центрального конца п. vagi, и твердой мозговой оболочки. Даже механическое раздражение последней с помощью губки с постоянством вызывало глотательные движения.
Здесь же считаем нужным сказать несколько слов и относительно полученных при наших опытах данных, относящихся до влияния мозговой коры на дыхание.
Как известно, вопрос о влиянии мозговой коры на дыхание Франсуа Франком разрешался таким образом, что «любой пункт
Рис. 2.
двигательной области мозговой коры при достаточно сильном раздражении и известной продолжительности последнего дает тот или другой эффект в изменении дыхательных движений»; другими словами, что «вся двигательная область оказывает равномерное влияние на дыхание». Однако большинство позднейших исследователей склоняется к тому взгляду, что для дыхательных движений имеются особые корковые центры.
Так, один из нас (проф. В. М. Бехтерев) в своем труде «Физиология двигательной области мозговой коры», начавшем выходить еще в 1886 г., между прочим замечает, что у всех животных с двигательной области мозговой коры удается вызвать резкий эффект на дыхание; не удавалось лишь точно определить положение точки, действующей на дыхательные мышцы у того или другого животного1. Позднее работами Мунка, Унферрих-та, Преображенского и др. на коре животных уже отмечаются центры, при раздражении которых получался резкий эффект на дыхание. Унферрихт, например, пишет, что «раздражение третьей извилины, по Геле, и второй, по Ландуа, у собак в одном месте, а именно кнаружи от центра для m. orbicularis, производит с постоянством замедление дыхания, причем продолжительность выдыхательной паузы удлиняется.
1 Бехтерев В. М., Физиология двигательной области мозговой коры, Архив психиатрии и пр. за 1886—1887 гг.
7*
99
Преображенский1 у кошек вызывал выдыхательную остановку дыхания, если раздражение производилось в области самого переднего и в то же время бокового или наружного отдела борозды, отделяющей вторую от третьей извилины; ему удалось определить также центр, с которого вызывался вдыхательный столбняк диафрагмы. У собак, однако, он не мог определить положение вдыхательного центра, хотя в отдельных случаях с различных мест он получал или ускорение, или замедление дыхания.
В наших опытах, производимых над собаками, мы получали резкое учащение дыхания с удлинением выдыхательной фазы каждый раз при раздражении самого переднего конца второйизвилины (по Ландуа), непосредственно над открытым нами центром глотательных движений. Это учащение дыхания тотчас же прекращалось, как только электроды отнимались от мозговой поверхности, и затем вновь возобновлялось вместе с приложением электродов к мозговой коре. В последнем случае нередко вслед за выдыхательной одышкой непосредственно следовал и глотательный акт, что само по себе служит доказательством ближайшего соседства центра для глотательных движений с центром, производящим выдыхательные движения.
1 Преображенский С, Ueber Atmungszentruminder Hirn-rinde, Wiener klin. Wschr., 1890, № 41—43.
К ВОПРОСУ О ВЛИЯНИИ МОЗГОВОЙ КОРЫ И ЗРИТЕЛЬНЫХ БУГРОВ НА ГЛОТАНИЕ
После того как наша общая с д-ром П. А. Останковым работа «О влиянии коры большого мозга на акт глотания и на дыхание» была сообщена в прошедшем 1893 году в одном из сентябрьских заседаний Казанского общества невропатологов и психиатров, мы получили возможность познакомиться с исследованиями Л. Рети *, который, производя опыты преимущественно на кроликах, нашел, что путем раздражения области, расположенной кпереди и кнаружи от коркового центра конечностей, вызываются жевательные движения, за которыми следует акт глотания.
По автору, эта последовательность движений обусловлена ничуть не тем обстоятельством, что второе движение рефлектор-но следует за первым, а тем, что эта последовательность движений уже дана в самой коре. При своих опытах над собаками мы также обратили внимание на ближайшее соседство в коре собаки центров глотания, центров движения челюстей, языка, углов рта, центров для открывания рта и т. п., но без специально поставленных опытов мы не можем пока утверждать, что уже в самой коре все вышеуказанные движения представляются сочетанными в одном и том же центре. Что же касается кроликов, то, повидимому, действительно имеется подобное сочетание, так как при раздражении коры у этих животных в вышеуказанной области глотательный акт обычно следует за жевательными движениями, центр которых на коре кролика был отмечаем уже и ранее некоторыми из авторов и, между прочим, мною в работе «Физиология двигательной области мозговой коры» 2.
Д-р Рети находит далее, что внутри или под thalamus opticus заложен сочетательный центр для еды. С отделением этого центра раздражение заложенных в ножке путей вызывает только отдельные сокращения жевательных мышц, без движений языка и губ и без глотательных движений.
1 Sitzb. d. К. Akademie d. Wissensch. in Wien, 1893, Bd. СП.
2 Архив психиатрии, нейрологии и судебной психопатологии за 1886 и 1887 гг.; отд. изд., стр. 24; см. также Arch, slaves de biologie за 1887 г.
101
В участии области зрительных бугров в жевательных и глотательных движениях у кроликов я сам убеждался неоднократно при опытах, производимых мною в прежнее время; при этом я считаю необходимым отметить здесь, что указание автора о содержании в зрительных буграх соче тательного центра для еды заслуживает большого внимания. Уже много лет, как я доказываю, что зрительные бугры содержат в себе центры для иннервации разнообразных растительных функций организма и для непроизвольных движений, участвующих в выражении ощущений1. Между прочим, в зрительных буграх, как показали мои опыты (при участии Н. А. Миславского), содержатся центры для движения желудка и кишок; само собой разумеется, что уже в силу этого обстоятельства представляется вероятной и зависимость иннервации пищевода от зрительных бугров, так как пищевод, как мы знаем, подобно желудку и тонким кишкам, ин-нервируется блуждающим нервом. Но исследования Рети ведут нас дальше: приняв их во внимание и руководясь полученными нами ранее данными относительно влияния зрительных бугров на желудочно-кишечную иннервацию, приходится признать, что путем раздражения зрительных бугров может быть приведена в деятельное состояние вся система движений, служащих для питательной функции организма, начиная с жевательных и глотательных движений и кончая движениями кишок.
1 Бехтерев В., Об отправлении зрительных бугров у человека и животных, Вестн. психиатрии и невропатологии, 1885; см. также Virch. Arch., 1887, Bd. 110. — Бехтерев и М и с л а в с к и й, О влиянии мозговой коры и центральных областей мозга на кровяное давление и деятельность сердца, Архив психиатрии, 1886; см. также Neurol. Zbl., 1886, № 18.—Мозговые центры движений мочевого пузыря, Архив психиатрии, 1887; см. также Neurol. Zbl., 1887. — О центральной и периферической иннервации кишок, Труды Общества естествоиспытателей при Казанском университете, т. XX; см. также A; h {. Anatomie und Physiologie, 1890.— О мозговых центрах движений влагалища у животных, Медицинское обозрение, 1891, № 15; см. также Arch. f. Anatomie und Physiologie, 1891.— Об иннервации и мозговых центрах слезоотделения, Медицинско; обозрение, 1891, № 12; см. также Neurol. Zbl., 1891.
О ВЛИЯНИИ МОЗГОВОЙ КОРЫ ЧЕЛОВЕКА
НА СЕРДЦЕБИЕНИЕ, ДАВЛЕНИЕ КРОВИ
И ДЫХАНИЕ
В литературе имеется уже целый ряд указаний относительно влияния мозговой коры на сердцебиение. Эти наблюдения сделаны на животных, которым производилось раздражение мозговой коры. Производя сам такого рода опыты у собак, я наблюдал учащение пульса, которое обыкновенно происходило вместе с поднятием кровяного давления при раздражении корковой области на месте gyri sygmoidei и соседних с ней областей. До сих пор, однако, не имелось наблюдений относительно подобных же влияний мозговой коры у человека. Ввиду этого мне кажутся не лишенными интереса наблюдения относительно влияния на сердцебиение поверхностных разрушений мозговой коры у человека, произведенных при операциях над мозговой корой.
В заведываемой мною клинике было произведено с 1895 г. три таких операции со вскрытием черепной покрышки в области корковых двигательных центров и с последующим удалением, вслед за вскрытием твердой оболочки, определенной части мозгового вещества. Во всех этих случаях, особенно же в двух случаях, вслед за операцией обнаруживалось крайне резкое учащение и полнота пульса, которое не могло быть объяснено ни состоянием температуры тела, остававшейся или на нормальной высоте, или же лишь короткое время державшейся на субфеб-рильном уровне, ни какими-либо иными поводами. Равным образом и со стороны дыхания у больных обнаруживались резкие изменения, которые должны быть поставлены в прямую зависимость от мозговой операции. Я приведу здесь и самые случаи, о которых идет речь.
Больной ребенок, 12 лет, с детства страдает тяжелыми и частыми эпилептическими приступами с судорогами, начинающимися с левой верхней конечности и поворачивания головы в левую сторону и затем быстро обобщающимися. 18 марта в 1 час дня ему сделана была в заведываемой мною клинике трепанация в области правых центральных извилин, причем по вскрытии мозга был удален центр поворачивания головы в противоположную сторону, обнаруженный мною с помощью фарадического тока при заднем отделе второй лобной извилины, и вместе с тем была удалена также часть центра для верхней конечности в среднем отделе передней центральной извилины. Вслед за тем твердая оболочка была зашита, костный лоскут вставлен на место, края кожной раны сшиты и на всю рану наложена
103
асептическая повязка. Утром того же дня с больным был небольшой припадок, состоявший в судорогах верхних и нижних конечностей, но без поворачивания головы. Затем до 1 мая, т. е. почти в течение около 2'/2 месяцев, припадков с больным не было вовсе. Впоследствии же они возобновились, но случались редко и под влиянием лечения Adonis vernalis с бромом даже и совсем прекратились на продолжительное время. Что касается его пульса, то он представлял крайнее учащение, вследствие чего после операции потребовалось назначение Inf. digitalis, который он принимал 21, 22 и 23 марта. Исследование пульса первоначально производилось три раза ежедневно в 9 часов утра, в 4 часа пополудни и в 8 или 9 часов вечера вместе с дыханием и температурой тела и дало следующие результаты.
|
|
9 часов утра |
4 часа пополудни |
9 часов вечера |
||||||
|
Дата |
пульс |
температура |
дыхание |
пульс |
температура |
дыхание |
пульс |
температура |
дыхание |
|
18 марта |
|
|
|
90 |
36,8 |
|
120 |
37,5 |
|
|
19 » |
120—136 |
37,6 |
— |
130 |
37,6 |
— |
126 |
37,8 |
— |
|
20 » |
110—120 |
37,0 |
— |
126 |
38,0 |
— |
■— |
— |
— |
|
21 » |
120 |
36,7 |
—. |
104 |
37,4 |
— |
108 |
37,3 |
— |
|
22 » |
106 |
37,1 |
— |
103 |
37,4 |
— |
108 |
37,4 |
— |
|
23 » |
96 |
36,7 |
— |
104 |
37,4 |
— |
102 |
37,2 |
- — |
|
24 » |
100 |
37,0 |
— |
106 |
36,9 |
28 |
90 |
37,1 |
28 |
|
25 » |
106 |
36,7 |
27 |
110 |
36,9 ' |
36 |
102 |
37,2 |
25 |
|
26 » |
104 |
36,7 |
24 |
108 |
37,2 |
30 |
102 |
37,3 |
28 |
|
27 » |
102 |
37,4 |
27 |
108 |
37,5 |
30 |
120 |
37,0 |
33 |
|
28 » |
104 |
87,2 |
26 |
114 |
37,3 |
33 |
90 |
37,1 |
24 |
|
29 » |
94 |
37,1 |
24 |
96 |
37,0 |
26 |
87 |
37,1 |
26 |
|
30 » |
96 |
36,9 |
30 |
90 |
37,1 |
27 |
94 |
37,0 |
30 |
|
31 » |
90 |
37,0 |
28 |
102 |
— |
28 |
80 |
37,2 |
24 |
|
1 апреля |
104 |
— |
30 |
100 |
— |
26 |
96 |
— |
241 |
|
2 » |
104 |
— |
30 |
96 |
— |
26 |
96 |
— |
24 |
|
3 » |
96 |
— |
24 |
86 |
— |
24 |
75 |
— |
26 |
|
4 » |
86 |
— |
24 |
90 |
— |
27 |
96 |
— |
27 |
|
5 » |
90 |
— |
27 |
88 |
__ |
24 |
90 |
—■ |
26 |
|
6 » |
80 |
— |
24 |
90 |
— |
27 |
84 |
—• |
24 |
|
7 » |
90 |
— |
27 |
80 |
— |
29 |
84 |
— |
26 |
|
8 » |
88 |
—. |
24 |
90 |
— |
27 |
90 |
—■ |
26 |
|
9 » |
90 |
— |
27 |
88 |
— |
26 |
75 |
— |
26 |
|
10 » |
86 |
— |
24 |
90 |
|
27 |
96 |
— |
27 |
|
11 » |
90 |
— |
27 |
60 |
|
24 |
84 |
— |
26 |
|
12 » |
86 |
— |
24 |
90 |
— |
27 |
96 |
— |
27 |
|
13 » |
90 |
—. |
27 |
86 |
— |
26 |
90 |
— |
26 |
|
14 » |
66 |
— |
24 |
86 |
— |
24 |
90 |
•— |
26 |
|
15 » |
90 |
— |
27 |
88 |
— |
21 |
90 |
— |
26 |
|
16 » |
90 |
— |
27 |
— |
— |
— |
94 |
— |
24 |
|
17 » |
|
|
|
|
|
|
|
|
|
|
18 » |
90 |
— |
27 |
— |
— |
— |
80 |
—■ |
24 |
|
19 » |
86 |
|
27 |
" |
~ |
|
80 |
|
26 |
1 Температура тела все остальное время держится с колебаниями от 36,8° Д0^37,3° и лишь однажды, а именно 2 апреля, поднялась до 37,5°.
04
Из этих данных нетрудно убедиться, что, несмотря на нормальную температуру тела, пульс у больного вслед за операцией представлял поразительное учащение, потребовавшее первоначально даже применения дигиталиса. Это учащение могло быть поставлено в связи исключительно с раздражением корковой поверхности, так как до операции пульс у больного, будучи неоднократно исследуем, не превышал е общем 80—86 ударов в 1 минуту.
Следует иметь в виду, что у нашего больного обращало на себя внимание то обстоятельство, что, несмотря на значительное учащение, пульс всегда представлялся полным и напряженным, что могло указывать на повышение бокового давления в сосудах.
Нельзя не отметить в этом случае и значительного учащения дыхания, наблюдавшегося вслед за операцией. Это учащение дыхания у больного может быть объяснено только тем, что корковый центр поворачивания головы был разрушен как раз в той области, в которой у животных, по моим исследованиям, произведенным совместно с Останковым1, помещается центр, раздражение которого вызывает резкое учащение дыхания. Отсюда очевидно, что и учащение дыхания, наблюдавшееся в вышеуказанном случае, следует рассматривать как результат раздражения мозговой коры в области дыхательного центра.
Перейдем теперь к рассмотрению другого случая, в котором было произведено повреждение мозговой коры на более обширном протяжении, но исключительно на центральных извилинах.
Больной, молодой субъект, с давних пор страдавший постоянными судорожными движениями правой половины тела, временами переходившими в настоящие падучие приступы, имевший рубцы на гглове хак следы бывших ушибов, был оперирован в заведываемой мною клинике проф. Вельяминовым в 1 час дня 4 февраля 1898 г. По остеопластическому способу была выдолблена лоскутообразная фигура в черепе соответственно левым центральным извилинам, затем нижняя стенка черепа надломлена и вместе с кожным лоскутом отворочена вниз. Вслед за тем твердая мозговая оболочка разрезана в форме подобного же лоскута и отворочена, после чего фарадическим током были определены мною центры для движения руки и лица. Под контролем фарадического тока эти центры затем были удалены, после того твердая оболочка была зашита, кость вложена на место и кожный лоскут сшит с краями раны. С самого момента операции пульс сильно участился, хотя температура держалась на нормальной или субфебрильной высоте и только однажды достигла 38,2°. Так, уже в день операции в 4 часа 15 минут пульс достиг 106 в 1 минуту при температуре 37°; в 5 часов 15 минут пульс 122 в 1 минуту; в 9 часов вечера—• 124 в 1 минуту, полный, дыхание 22, температура 37,7°; в 1 час 16 минут ночи пульс 128, полный, дыхание 36 при температуре 38,2°; в 2 часа 45 минут пульс 100 в 1 минуту, полный, дыхание 24 в 1 минуту, ровное, температура 37,9°; в 5 часов утра пульс 100 в 1 минуту, температура 37,6°; в 7 часов утра пульс 125 в 1 минуту, полный и значительно напряженный, дыхание 22, температура 37,7°.
1 Неврологический вестник, 1893; Neurol. Zbl., 1893; см. также сообщение доктора Жуковского (из моей лаборатории) в научном собрании врачей клиники душевных и нервных болезней за 1897 г.
105
На другой день, 5 февраля, в 9 часов утра пульс 120 в 1 минуту, полный, дыхание 20 в 1 минуту, температура 37,4 ; в 4 часа пополудни пульс 134 в 1 минуту, температура 37,2°; в 9 часов вечера пульс 108 в 1 минуту; 6 февраля в 11 часов утра пульс 108 в 1 минуту, вечером около' 100 в 1 минуту; 7 февраля пульс учащен, но не был сосчитан; 8 февраля пульс 86 в 1 минуту; 10 февраля утром пульс 108 в 1 минуту; 11 февраля пульс 108, температура 36,1°. 13 февраля с больным было 3 судорожных приступа, после чего наступил продолжительный сон.
14 февраля в 12 часов дня пульс 84 в 1 минуту, температура 37,5°. После того был судорожный приступ; в 3 часа дня пульс 72 в 1 минуту, полный, дыхание 24 в 1 минуту.
jib 15 февраля к 12-му часу с больным случился судорожный приступ, после которого пульс 106 в 1 минуту, аритмичный, температура 36,5°; 16 февраля пульс 86, температура 37°.
Остальные дни пульс представлялся нормальным, около 72—84 в 1 минуту, обыкновенного напряжения.
Таким образом, в приведенном случае пульс тотчас же вслед за операцией удаления мозговой коры достиг 106 в 1 минуту и затем перешел к вечеру того же дня за 120 в 1 минуту при нормальной и субфебрильной температуре тела. Также и на другой день вслед за операцией пульс достиг 120 в 1 минуту и даже 134 в 1 минуту при температуре 37,2—37,4°; остальные же дни до 13—14 февраля, следовательно, в течение около 9—10 дней вслед за операцией, держался выше 100 при нормальной температуре тела. Достойно внимания, что и в этом случае пульс, несмотря на частоту, представлялся вслед за операцией все время полным и напряженным, что указывает на повышенное давление крови.
Что касается дыхания, то здесь не обнаруживалось резкого учащения, как в первом случае, что вполне гармонирует с тем обстоятельством, что разрушение ограничивалось исключительно областью центральных извилин, следовательно, было произведено позади дыхательного центра.
Оба вышеприведенные наблюдения, таким образом, не оставляют сомнения в том, что двигательные области мозговой коры у человека, как и у животных, обнаруживают влияние на сердцебиение, давление крови и дыхание.
Что касается собственно центров сердцебиения и давления крови, то, имея в виду, что в обоих вышеприведенных случаях разрушение было произведено на различных участках двигательной области мозговой поверхности, необходимо притти к выводу, что и у человека, подобно тому, как у животных, центры, управляющие движением сердца и давлением крови, раскинуты на значительном протяжении двигательной площади мозговой коры и соседних с нею областей. Что же касается центра для дыхания, то у человека он, как и у животных, очевидно, располагается в передних отделах двигательной площади, по всей вероятности, на заднем отделе второй лобной извилины, как показывает первое из наших наблюдений.
ВЛИЯНИЕ КОРЫ ГОЛОВНОГО МОЗГА НА ПОЛОВЫЕ ОРГАНЫ, ПРЕДСТАТЕЛЬНУЮ И МОЛОЧНЫЕ ЖЕЛЕЗЫ1
Несмотря на огромную роль, которую играют психические факторы в половой деятельности, вопрос о влиянии коры головного мозга на состояние половых органов остается совершенно недостаточно освещенным в литературе.
Поскольку мне известно, впервые мною совместно с д-ром Н. Л. Миславским в 1890—1891 гг. был поставлен ряд опытов на кроликах и собаках для выяснения влияния коры головного мозга на движения влагалища2. При этом мы применяли тогда во всех опытах графический метод регистрации. Тонкий резиновый баллон вводился во влагалище либо через половую щель, либо через один из рогов матки. Он наполнялся водой и соединялся с водяным манометром, колебания которого записывались при помощи капсулы Марея на кимографе.
В этих опытах мы убедились в том, что путем соответствующих раздражений коры головного мозга можно вызвать или затормозить движения влагалища.
У кролика область коры головного мозга, раздражение которой вызывает известные изменения состояния влагалища, соответствует полям, в которых расположены моторные центры. Возбуждающие влияния на движения влагалища у этого животного проявляются в более или менее выраженном ускорении и повышении нормальных сокращений, тормозящее влияние — в виде замедления этих сокращений и даже более или менее длительном их прекращении.
У собаки раздражение коры головного мозга оказывало значительно менее выраженное влияние на движения влагалища, чем у кролика. Оно вызывалось также со стороны gyrus sigmoi-deus. Лишь в немногих опытах удалось при увеличении силы тока вызвать эффект со стороны влагалища при раздражении точек, расположенных возле gyrus sigmoideus кнаружи или кну-
1 Archiv fur Anatomie und Physiologie, Physiologische Abteilung, 1905, S. 504—537.
2 W. Bechterew und N. Mislawski, Ueber die Hirn-zentren der Scheidenbewegungen bei Tieren, Arch. f. Anatomie und Physiologie, Phisiol. Abt., S. 380.
107
три от него. У собаки, так же как у кролика, со стороны gyrus sigmoideus можно вызывать как возбуждающий, так и тормозящий эффект. В первом случае он проявлялся в более или менее выраженном усилении нормальных движений влагалища, во втором случае раздражение коры обусловливало замедление и ослабление, вплоть до полного прекращения движений влагалища на некоторое время.
Увеличение сокращений влагалища под влиянием раздражений коры головного мозга выражается обычно в виде довольно высокой и длинной волны сокращения, то появляющейся, то исчезающей и имеющей даже несколько вершин неравной высоты. В дальнейшем, после того как нанесение раздражений прекращалось, обычно наступало удлинение и ослабление сокращений влагалища, иногда даже их полное исчезновение.
В случае тормозного эффекта проявления были иного характера. Сначала при нанесении раздражений сокращения влагалища становились слабее и медленнее. Далее они вовсе прекращались, причем в большинстве случаев на все время, пока наносились раздражения. После прекращения раздражений обычно наступало значительное усиление сокращений, выражавшееся в виде волны с рядом вершин или в виде ряда волн различной высоты и длительности.
В некоторых наших опытах после периода полного прекращения сокращений влагалища наступал настоящий тетанус влагалища в форме сильного, длительного сокращения.
Если раздражение коры, оказывающее тормозящее влияние, наносилось в момент сокращений влагалища, то последние моментально уменьшались и даже полностью прекращались. По прекращении раздражений, как обычно, наступали усиленные сокращения.
Коротко —явления, обнаруженные при раздражении коры головного мозга, с несомненностью показывают, что влияние коры головного мозга на движения влагалища осуществляете? посредством локального нервного механизма, поддерживающего определенный мышечный тонус влагалища и его самостоятельные периодические сокращения. Если раздражения наносятся слишком длительное время, то возникающее усиленное возбуждение этих механизмов приводит, вследствие утомления, в конечном счете к преобладанию тормозного эффекта. Приостановка сокращений под влиянием кортикальных импульсов приводит к преобладанию сил, вызывающих процесс возбуждения, что ведет к усилению и ускорению сокращений.
Относительно локализации полей, оказывающих тормозящее или возбуждающее влияние, из наших опытов следует, что возбуждение движений влагалища с последующим их торможением наиболее постоянно возникает при раздражении задней части gyrus sygmoideus, за исключением его боковой части, и не столько-постоянно при раздражении наиболее медиальной части перед-
108
него отдела извилины. Наоборот, торможение движений влагалища с последующим усилением движений в наших опытах вызывалось чаще всего раздражением латеральной части заднего отдела gyrus sigmoideus, области, переходной от задней к передней части, а также наружно-передней части gyrus sigmoideus.
В отношении топографии действующих корковых полей существуют значительные индивидуальные особенности. В большой мере эффект, помимо того, зависит также от возбудимости коры головного мозга: иногда при раздражении одной и той же точки коры головного мозга то получают соответствующий эффект в виде возбуждения, то не получают, а иногда даже возникает противоположный эффект. Это свидетельствует о том, что в коре головного мозга нет, собственно говоря, строго диференцирован-ных тормозных и возбуждающих полей. Можно полагать, что в коре существует лишь активная область, связанная с движениями влагалища, из которой могут исходить импульсы как возбуждающие, так и тормозящие движения влагалища. При этом в нормальных условиях при определенной силе раздражителей из одних пунктов вызывается преимущественно возбуждающий, из других —тормозной эффект. При изменении возбудимости поля или силы тока вместо одного эффекта может возникать противоположный.
В отличие от сокращений влагалища, наступающих под влиянием раздражения спинного, продолговатого мозга и зрительного бугра, при раздражении коры наступает не однократное длительное сокращение, а либо волна сокращения значительной длины и высоты, имеющая ряд вершин, либо ряд сильных, быстро следующих друг за другом сокращений. При этом обнаруживается отчетливый латентный период; это также говорит в пользу того, что с одной и той же точки могут возникать как возбуждающие, так и тормозящие влияния и что наступающий эффект зависит от преобладания то одних, то других.
Таким образом, в период времени, соответствующий латентному периоду, происходит как бы борьба между тормозящими и возбуждающими влияниями. Если одни из них начинают преобладать, то наступает более или менее значительное колеблющееся в своей интенсивности изменение кривой.
В связи с вопросом о влиянии коры головного мозга на двигательные способности влагалища стоит вопрос о влиянии коры головного мозга на движения матки.
По этому вопросу до последнего времени имелись лишь весьма неопределенные данные. Некоторые авторы вообще отрицали возможность влияния коры головного мозга на движения матки.
Хотя Гедус еще в 1852 г. во время одного опыта наблюдал сокращения матки при раздражении коры головного мозга, однако эти опыты оставались в литературе незамеченными, и
109
позднейшие авторы пришли вообще к отрицанию возможности влияния коры на движения матки.
По Гауху1, движения матки наступают при раздражении мозжечка, продолговатого мозга и всего спинного мозга, особенно его поясничного и крестцового отделов, в то время как раздражения коры головного мозга в его опытах не приводили к появлению каких-либо движений матки. Наоборот, раздражение глубоких частей полушарий обычно вызывало появление сокращений матки.
Кроме того, в литературе имеются отдельные замечания у Бошфонтэна, которому удалось в своих опытах вызвать сокращения матки путем раздражения gyrus sigmoideus. Сам Бошфон-тэн пытался объяснить эти явления рефлекторными влияниями.
Влияние коры головного мозга на движения матки подтверждается также клиническим наблюдением.
Известно, что сильные аффекты, как испуг, радость и т. п.„ могут резко усилить родовую деятельность и даже привести к аборту или преждевременным родам.
С другой стороны, общеизвестно также тормозящее влияние психических факторов на родовой акт. Появление незнакомого человека в родильной комнате приводит иногда к моментальному уменьшению или полному прекращению схваток. Это нельзя иначе объяснить, как торможением движений матки, исходящим со стороны корковых центров.
Для того чтобы проследить влияния коры головного мозга на движения матки, по моему предложению доктором Плохин-ским2 в нашей лаборатории были поставлены опыты на кроликах с применением графической регистрации.
После вскрытия брюшной полости и выведения рогов матки наружу в один из рогов вставлялся маленький резиновый баллончик. Для этого производился небольшой разрез по овариаль-ному концу рога. Брался катетер, на один конец которого надевался маленький баллон; второй конец катетера оставался свободным. Этот свободный конец катетера выводился через разрез в роге матки, шейку матки, влагалище и половую щель наружу настолько, чтобы баллон на другом конце катетера оказался в полости матки. Введенный в полость матки баллон заполнялся водой, после чего катетер соединялся с манометром. Колебания манометра, т. е. объема баллона, записывались на барабане. Сокращения матки, естественно, уменьшали объем баллона и вызывали подъем кривой, расслабления матки позволяли баллону расширяться, в связи с чем кривая падала вниз (аналогично тому, что имело место в вышеуказанных опытах с регистрацией движений влагалища).
1 Н а и с h, Ueber den Einfluss des Riickenmarks und Gehirns auf die Bewegungen des Uterus. Inaug. Dissertation, Halle, 1879.
2 Плохинский, Обзор психиатрии, СПБ, 1902.
110
Для опытов брались обычно либо беременные, либо в прошлом рожавшие животные. В первом случае баллон вводился непосредственно в рог, без введения катетера в матку.
Все опыты ставились без наркоза, под местной анестезией 2% раствором кокаина.
Наши опыты показали, что медиальная сторона задней части gyrus sigmoideus, т. е. моторное корковое поле, является той действенной областью, из которой могут быть получены сокращения матки. Иногда это место находится в задней части моторной зоны на внутреннем крае полушария, иногда лежит более кнаружи.
Если уже были спонтанные сокращения матки, то раздражение этого места вызывало усиление сокращений.
Если же во время опыта спонтанных сокращений матки не было, то раздражение вышеуказанных мест коры нередко вызывало ряд ритмических сокращений. Изредка в этих случаях наблюдалось единичное сокращение, но и тогда при повторных раздражениях вызывался ряд ритмических сокращений.
В некоторых случаях многократные раздражения вызывали все более и более сильные сокращения матки. С прекращением раздражений сокращения матки становились обычно слабее и в конце концов полностью прекращались. Если кора вновь подвергалась раздражению, то в некоторых случаях удавалось вновь вызвать ритмические сокращения матки. Возбудимость центров при этом в процессе опыта все более и более падала.
Действие центров матки является двусторонним, так как из каждого полушария можно получить одинаковый эффект.
Латентный период в этих случаях, так же как и в случаях вызывания сокращений влагалища путем раздражения коры, крайне длительный —от 10 до 16, 30 и даже 45 секунд.
Из отдельных опытов следует, что кора головного мозга может являться источником тормозных влияний на ритм сокращений матки.
В этом случае мы также можем различать тормозящее и возбуждающее действие. При этом первое выражается в появлении новых или усилении уже имевшихся сокращений матки, второе — в ослаблении или полном подавлении ритмических сокращений.
Ритмика маточных сокращений, зависящая от автоматической деятельности ганглиев матки, также находится под известным влиянием корковых центров. Уже в обычных условиях раздражения, идущие от центральных органов, могут оказывать то усиление и ускорение, то уменьшение и замедление сокращений матки. Этим объясняется то, что после раздражения корковых центров, так же как и центров в зрительном бугре, влияние которых установлено вышеуказанными исследованиями, ритм нарастает или вновь выступает, в то время как в других случаях он более или менее подавляется или вовсе приостанавливается.
Ш
Что касается длительности латентного периода сокращений матки при раздражении коры, то она объясняется, вероятно, тем, что под влиянием раздражения возникают как возбуждающие, так и тормозящие влияния, и что при этом в конце концов начинает преобладать возбуждающее действие.
Влияние коры головного мозга на сокращение матки может быть обнаружено и у собак, но в общем у них оно менее выражено, чем у грызунов.
Во время эпилептических припадков у собак постоянно отмечается появление или повышение сокращений матки. При этом непрерывных тетанических сокращений не наблюдается. Последние носят периодический характер и имеют обычную длительность.
Так как активность сокращений матки зависит, как вытекает из соответствующих исследований, не только от гладких мышечных волокон, но и от соответствующей величины притока крови, корковые центры, влияющие на сокращения матки, должны оказывать влияние и на физиологические процессы, именуемые менструациями. В этом отношении нет специальных экспериментальных исследований, но клинический опыт с несомненностью указывает на эту связь.
Менструации могут под влиянием чийто психических воздействий, страха и т. п. внезапно прекратиться вследствие тормозных влияний, исходящих от мозга. С другой стороны, известно, что психические моменты могут оказывать и противоположное влияние —вызывать преждевременное наступление менструаций, что, несомненно, стоит в связи с возбуждающим действием на менструации импульсов, идущих из мозга.
Исследования в области гипнотизма и внушения показали, что даже обильные маточные кровотечения могут быть приостановлены путем гипнотического внушения, как мне приходилось неоднократно наблюдать1.
Разумеется, в коре головного мозга должны быть центры, оказывающие влияние на мужской половой аппарат. В этом нельзя сомневаться хотя бы потому, что психические факторы оказывают огромное влияние на половую сферу. Так называемое половое влечение по своей природе является психорефлекторным процессом.
Как показывают опыты Гольца, удаление полушарий у самца лягушки сопровождается утратой им полового влечения к самке. Даже в присутствии последней оперированный самец остается индиферентным. Из этого следует, что половое влечение связано с полушариями, которые оказывают на него влияние под дей-
1 В. М. Бехтерев, О сдавлении поясничных центров спинного мозга и др.; Значение гипноза в качестве лечебного средства, в кн.: Нервные болезни в отдельных наблюдениях, Казань, 1884.
В. М. Бехтерев, Терапевтическое значение гипноза, СПБ.
112
ствием внешних раздражителей. Более низшие центры в той или иной степени подчинены высшим.
В животном царстве в качестве возбудителя полового влечения особенную роль из органов чувств играет обоняние.
Обоняние у большинства млекопитающих имеет несравнимо большее значение для возбуждения половой функции, чем зрение.
Что касается влияния коры головного мозга на эрекцию полового члена, то оно весьма вероятно еще и потому, что сокращения мышц, участвующих в акте совокупления, в известной мере подчинены воле. С другой стороны, п. errigens возбуждается не только рефлекторным путем, но и под влиянием соответствующих представлений.
Специальные исследования устраняют всякие сомнения относительно существования особого кортикального центра эрекций полового члена.
Я сам во время опытов на собаках имел возможность убедиться в том, что раздражение задней доли gyrus sigmoideus в месте, соответствующем расположению центров влагалища у самки, вызывает у самца напряжение и увеличение в размерах полового члена.
Исчерпывающее исследование коркового эрекционного центра у собак с применением графического метода было, по моему предложению, проведено в нашей лаборатории доктором Пуссепом1.
Регистрация напряжений penis производилась с помощью охватывающего его наполненного водой сосуда, соединенного резиновым рукавом с манометром, а через него —с мареевским барабанчиком.
Из этих исследований вытекает, что раздражение упомянутого поля в заднем отрезке gyrus sigmoideus вначале влечет за собой некоторое снижение кривой, что соответствует уменьшению объема члена, а затем большее или меньшее повышение кривой, соответствующее наступившему напряжению и увеличению объема члена.
Что касается начального падения кривой, то оно может зависеть от сокращения относящихся к penis мышц или от сокращения сосудов penis.
Для решения этого вопроса имело смысл произвести исследования с кураре, который парализует произвольные мышцы. Проведенные опыты показывают, что кривая в этих случаях обнаруживала значительно меньшее падение, но и дальнейший подъем кривой был тоже меньше, чем без применения кураре. Другими словами, кураре ослаблял эрекцию penis в большей или меньшей степени. Так как и прямое наблюдение не указывало на зависимость падения кривой от состояния мускулатуры penis, то следовало отнести начальное падение кривой за счет сужения сосудов. Этот факт приводит к заключению, что
1 П у с с е п , InaUg. Dissertation, СПБ, 1902.
8 В . М. Бехтерев
113
внешние раздражения коры имеют также тормозящее действие на половой член.
Дальнейшее исследование показало, что при раздражении коркового центра током слабым или средней силы (от 10 до 1? К—А) начальное падение кривой менее сильно и продолжительно, чем в случаях раздражения более сильным током (от 6 до 8 К—А). В то же время обнаружилось, что в упомянутой области коры имеются два поля, из которых верхнее или внутреннее при умеренном раздражении всегда вызывает более длительное падение кривой, наряду с тем, как раздражение нижнего или наружного поля влечет сравнительно больший подъем кривой. Таким образом, в первом поле сконцентрированы сосудосуживающие элементы, во втором —сосудорасширяющие. Но оба поля вызывали в итоге одинаковое напряжение полового члена при значительном отклонении интенсивности и длительности первого, т. е. сосудосуживающего эффекта.
Здесь, как и при влиянии мозговой коры на движения влагалища, также видно, что возбуждающее и тормозящее действие проявляются не в чистом виде, а более или менее смешанными.
Исследование состояния сосудов penis способом, подобным методу измерения давления крови в мозгу по Гюртле, приводит к аналогичным данным: раздражение нижнего поля вначале дает меньшее сужение сосудов и впоследствии большее расширение их, в то время как раздражение верхнего поля обусловливает сужение сосудов.
Еще поучительнее были данные, полученные при одновременном исследовании давления в v. dorsalis penis ива. cruralis, причем выяснилось, что раздражение нижнего поля очень повышает давление в вене, общее же давление в артерии голени мало изменяется; в то же время раздражение верхнего поля влечет за собой незначительное повышение давления в вене.
Отмечено, что оба полушария оказывают приблизительно одинаковое действие на penis, из чего проистекает, что центр эрекции полового члена —двусторонний.
При удалении у собаки самца центра эрекции из обоих полушарий он по исчезновении операционного шока не обнаруживает склонности приближаться к самке и во время течки, хотя механическая возбудимость полового члена у таких собак даже повышена. Собаки с подобными повреждениями головы и мозга, но в других областях коры обнаруживают такое же половое влечение, как и во время здоровья. При более сильном раздражении центра эрекции, именно его нижнего отрезка, наступает вместе с напряжением полового члена и эякуляция семени, причем можно наблюдать даже соответствующие coitus движения задних конечностей.
Поскольку особый центр эякуляции в коре не найден, надо считать, что центр эрекции одновременно представляет и эякуляцию.
114
Из упомянутых прежде исследований вытекало также, что если половой член эрегирован путем раздражения спинального центра, то раздражение коркового центра еще повышает возбуждение его. Ясно, что кортикальные, т. е. психические, импульсы не только не понижают рефлекторное возбуждение полового члена, но еще и усиливают его.
Раздражение обонятельной доли не имеет влияния на эрекцию полового члена, как показывают аналогичные опыты. Двустороннее удаление обонятельных долей также не нарушает половых функций у собаки. Подобным образом оперированные животные обнаруживают такое же libido, как здоровые, с той лишь разницей, что, не имея возможности пользоваться обонянием, они удовлетворяют свое половое влечение не только с самками в периоде течки, но и с теми, которые не находятся в этом состоянии.
Утверждение многих авторов, что обоняние играет главную роль в половом влечении, является,_ видимо, неприемлемым.
Можно считать, что обоняние играет роль важного вспомогательного момента при возбуждении полового влечения у некоторых животных, но не больше, так как при недостатке обоняния у этих же животных возможно удовлетворение полового влечения.
Кроме обонятельных впечатлений, для возбуждения полового влечения некоторое значение имеют зрительные, осязательные и даже слуховые воздействия (пение, музыка); у животных (и у извращенных людей) —также вкусовые ощущения. Но во всех этих случаях речь, очевидно, идет о возбуждениях, которые освобождают и поддерживают половое влечение, в существенных же своих проявлениях половое влечение, видимо, подчинено упомянутым центрам мозговой коры.
Далее, не подлежит сомнению, что секреция половых органов также находится в зависимости от коры больших полушарий. Значение психики для выделения спермы настолько известно, что нет. необходимости говорить об этом. Однако в последнее время не имелось, насколько мне известно, исследований о зависимости секреции спермы от мозговой коры. В моей лаборатории был в этом направлении предпринят вновь ряд исследований, осуществленных, по моему предложению, доктором Пус-сепом.
Методически речь шла о введении тончайшей канюли в семя-выводящие протоки с обнажением яичка.
Опыты показали, что раздражение коркового поля, расположенного по соседству с центром, возбуждающим эрекцию, и частично ему идентичного, вызывало повышенное выделение семени наряду с видимым простым глазом расширением сосудов яичка. Действие всегда выявлялось на стороне раздражения. Совсем иные явления наблюдаются по соседству с центром, затормаживающим эрекцию. Здесь сосуды яичка той же стороны
8*
115
обнаруживают вьпаженное сокращение. Предположение, что это действие должно совпадать с уменьшением секреции спермы, вполне возможно, но точно проследить это не удается, так как секреция семени вообще очень незначительна.
Для того чтобы точно установить, не является ли повышенное выделение семени при раздражении окружности возбуждающего эрекцию центра следствием какой-либо механическойпричины, например, результатом сокращения семенных канальцев, в опытах вводился атропин. Выявилось, что, несмотря на это, при раздражении коркового центра секреция семени увеличивалась. О зависимости раздражающего действия от мускулатуры яичек здесь не может быть речи, так как во время опытов яички были отпрепарированы и высвобождены.
Действие коры, по крайней мере в этих случаях, должно быть отнесено частично к повышению секреции семени, а не только к простому внешнему выражению. Что касается вопроса, имеем ли мы здесь дело с секреторным эффектом в собственном смысле или с вазомоторным действием,, которое также ведет к повышению выделения спермы, то он еще не может быть полностью разрешен. Во всяком случае, из проведенных опытов, несомненно, вытекает, что в мозговой коре в области двигательной зоны имеется истинный центр секреции спермы, возбуждающий деятельность семенных желез.
Исследованиями, проведенными в моей лаборатории, открыт кортикальный центр секреции предстательной железы по соседству с центром секреции спермы.
Исследования относительно локализации этого центра принадлежат доктору Пуссепу. Все опыты проведены на собаках в возрасте от 2 до 4 лет, когда предстательная железа особенно активна; в большинстве опытов применялся кураре. Требовалось устранить признанное в более ранней методике неудобным введение канюли в уретру, так как при этом секрет железы мог смешаться с выделениями мочевого канала. С этой целью подопытному животному производится разрез по средней линии под симфизом, причем после отделения околопузырной клетчатки обнажались шейка пузыря и предстательная железа. Затем мочеточник перевязывался ниже простаты, также и семенники; разрезался мочевой пузырь и в него вводилась канюля, которая прикреплялась к cervix vesicae швом, наложенным между слоями пузыря, во избежание повреждения нервов. Канюля соединялась с тонкой трубкой, после чего количество секрета устанавливалось путем счета вытекающих капель. В другой разрез вводилась канюля, соединенная с резиновой трубкой.
Организованные таким образом опыты привели к следующему результату: раздражение маленького поля, расположенного приблизительно на 0,5 см кзади от sulcus cruciatus и отстоящего примерно на 1 см от большой продольной борозды мозга, вызывало всегда повышенное выделение секрета предстательной
И6
железы. В общем это поле находится кзади и несколько книзу от места, которое оказывает влияние на выделение семени.
Здесь, так же как и в других случаях, раздражение коры сопровождается значительно большим скрытым периодом, чем раздражение тех спинальных и подкорковых областей, которые воздействуют на секрецию предстательной железы. Применение более сильного тока от 4 до 5 К—А прекращало выделение секрета, видимо, вследствие тормозящего действия. В некоторых опытах создавалось впечатление, что кора книзу и позади от упомянутого места оказывала в известной мере тормозящее влияние на секрецию prostatae, но это положение требует еще проверки дальнейшими исследованиями.
С половым аппаратом тесно связана служащая для питания ребенка деятельность молочной железы.
Гладкая мускулатура грудных сосков, способствующая выделению молока, также находится под явным воздействием корковых импульсов, за что говорит уже связь с психическими моментами. Известно, что при некоторых психических состояниях молоко струей истекает из сосков. Трудно сказать, идет ли здесь речь о мускульном влиянии или о собственно секреторных процессах. Возможны оба эти случая. Однако ясно, что мозговая кора должна иметь особый центр для грудной железы. Никто не сомневается в том, что отделение молока в некоторой степени находится под влиянием коры, так как известно, что психические состояния, сопровождающиеся приятным чувством, способствуют набуханию грудных желез и вызывают обильное выделение молока; в то же время противоположные эффекты уменьшают секрецию молока. Внезапные психические потрясения, как страх и т. п., прекращают секрецию молока.
При виде плачущего голодного ребенка грудные железы многих женщин явственно набухают. Известно также, что коровы дают больше молока, когда они кормят теленка, в то время как беспокойство животного или доение чужими руками уменьшают количество молока.
Бушю сообщает о случае, когда у кормящей женщины сразу пропало молоко при виде того, как ее ребенок упал; молоко вновь появилось, когда ребенок потребовал грудь.
В литературе имеются наблюдения, из которых вытекает, что не только количество, но и качество молока может изменяться в связи с психическими моментами, особенно при состояниях душевного угнетения. Бордо рассказывает, что молоко у кормилицы свернулось. Известны случаи, когда молоко при печальном настроении становилось кислым, причем у ребенка возникли эпилептические припадки (Месли). Берлин видел эклампти-ческие судороги у младенца, которому дала грудь раздраженная женщина. Левре наблюдал такие судороги у щенка после того, как животное сосало грудь женщины, находившейся в гневном возбуждении.
117
К сожалению, во всех этих случаях, большинство из которых относится к более давнему времени, не было произведено химическое исследование молока. Тем не менее изложенное не оставляет сомнения в том, что психические моменты имеют большое влияние на количество и, вероятно, также на качество молока. Фогель был частично прав, когда сравнивал секрецию молока в ее зависимости от аффективных состояний психики с выделением слез.
К сожалению, нет еще специальных исследований корковых областей, воздействующих на секрецию молока.
Однако, поскольку деятельность грудной железы, как и всех других желез, должна находиться под вазомоторным влиянием со стороны нервной системы и поскольку мы знаем, что вазомоторные центры для различных областей тела следует искать в большом чувствительном поле мозговой коры, можно принять, что в сензо-моторной области могут располагаться и центры, способствующие повышенному выделению молока. Этот вопрос должен быть разрешен лишь специальными экспериментальными исследованиями.
Такие исследования по моему предложению были проведены недавно в нашей лаборатории доктором Никитиным *.
Для опыта использовались овцы в периоде лактации. В молочные железы им вводилась стеклянная канюля и количество молока определялось или простым счетом вытекавших из канюли капель, или с помощью особого регистрирующего прибора, который отмечал падавшие капли на вертящемся барабане. После введения канюли выжидали время до прекращения спонтанного выделения молока. Всего вместе с предварительными опытами было проведено 28 исследований с раздражением коры. При этом оказалось, что секреция молока, хотя и не с очень большим постоянством (в 5 из 28 опытов), могла быть вызвана раздражением двигательной зоны мозговой коры по соседству с лицевым центром (область губ).
Обычно секреция появляется после длительного скрытого периода, причем сильнее из противоположной железы, чем из расположенной на одноименной стороне, хотя и последняя тоже увеличивает свою деятельность при раздражении коры. Иногда можно было наблюдать и consecutive последовательное выделение молока.
Заслуживает внимания, что более поздние периоды лактации не благоприятствуют корковому влиянию на секрецию молока.
Состав молока при этих опытах исследовался, но полученные результаты еще не могут считаться окончательными. Предстоит также провести опыты с одновременным измерением кро-
1 Никитин, О влиянии мозга на секрецию молока. Сообщение на научном заседании психиатрической и нервной клиник в С.-Петербурге, март 1905.
118
вяного давления. Раздражение затылочных и височных долей давало в опытах отрицательный результат.
У овец могут быть прослежены также влияния психических и других раздражений на выделение молока. Вспышки магния обусловливали уменьшение и даже прекращение выделения молока, выстрелы из пистолета и болевые раздражения тоже вызывали его исчезновение.
Таким образом, проведенные опыты не оставляют сомнения в том, что в сензо-моторной зоне мозговой коры расположены центры, раздражение которых вызывает отчетливые изменения секреции молока. Эти центры имеют отношение к приведенным выше наблюдениям, говорящим о возможности психического влияния на секрецию молока.
О ЗНАЧЕНИИ ОРГАНОВ РАВНОВЕСИЯ
В ОТНОШЕНИИ РАЗВИТИЯ НАШИХ ПРЕДСТАВЛЕНИЙ
О ПРОСТРАНСТВЕ1
Уже с самых первых своих шагов в исследовании физиологической роли органов равновесия, результатом которых явился ряд моих статей, помещенных в различных русских журналах и в «Archiv fur die gesamte Physiologie» 2, я пришел к выводу о важном значении этих органов в образовании наших представлений о пространстве. В 1884 г. в «Вестнике судебной и кли-
1 Настоящая статья представляет собою собственно переработку статьи, помещенной в Вестнике психиатрии за 1884 г. и вышедшей в том же году отдельным изданием под заглавием «Теория образования наших представлений о пространстве». Первоначально она была приготовлена мною собственно для иностранной печати, в которой я до сих пор еще не имел возможности заявить о своих взглядах в указанном отношении. Так как, однако, со времени обнародования вышеуказанной статьи иа рус-ком языке истекло уже более десятка лет, то естественно, что за такой период времени не могло не произойти некоторого изменения взглядов в теоретических воззрениях на указанный предмет, а следовательно, и в отношении тех или других частностей в объяснениях явлений, наблюдаемых при поражении органов равновесия. Естественно поэтому, что и переработка вышеуказанной статьи, не говоря о некоторых дополнениях, основанных на позднейших научных данных, оказалась незначительной, в силу чего я счел за правильное напечать настоящую статью не только на немецком языке, но и на русском, тем более, что упомянутое выше отдельное издание работы на русском языке давно уже вышло из продажи.
2 См. мои статьи: «Исследования вынужденных или насильственных движений (Zwangsbewegungen) у животных при разрушении стенок воронкообразной части третьего желудочка в мозгу», Еженедельная клиническая газета, 1892; см. также труды Общества русских врачей за 1882 г. — «Tierversuche iiber zwangsweise Rollbewegungen um diej Langsachse», St.-Petersb. med. Wschr., 1882; «Die Bedeutung der Trichterregion des 3-ten Venlrikels fur die Erhaltung des Korpergleichgewichtes», St.-Petersb. med. Wschtr., 1882 — «К физиологии равновесия тела. Отправление центрального серого вещества третьего мозгового желудочка», Военно-медицинский журнал, 1883; Pfliiger's Arch., 1883, Bd. XXXI.—«Об отправлении оливчатых тел продолговатого мозга», Врач, 1882, № 35; Pflii-ger'sArch., 1882, Bd. XXIX.—«Опыты над перерезкой слухового нерва у собак», Протоколы заседаний Общества психиатров за 1882 г.; Труды Общества русских врачей, 1882.—«Об отправлении полукружных каналов перепончатого лабиринта»,-Международная библиотека, 1882; Pfliiger's Arch., 1882, Bd. XXX. —«О связи так называемых периферических органов равновесия с мозжечком», Русская медицина, 1884, 3; Pfliiger's Arch., 1884, Bd. XXXIV.
120
нической психиатрии» я посвятил этому предмету специальную работу, в которой старался выяснить свои взгляды в этом отношении \ Так как, однако, вопрос о роли органов равновесия в образовании пространственных представлений является в высокой степени важным в научном отношении и его лишь изредка касаются в физиологической литературе последнего времени, то я позволю себе в этой статье отчасти воспроизвести в сокращенном виде, отчасти развить те отделы только что указанной работы, которые и по настоящее время, на мой взгляд, сохраняют свою научную ценность и интерес; все же остальное да позволено мне будет или опустить или изменить соответственно научным данным новейшего времени.:
Продолжительные экспериментальные исследования, предпринятые мною относительно так называемых органов равновесия, привели меня к тому заключению, что эти органы не только-служат для целей равновесия, но и доводят до сознания определенные ощущения, характер которых до сих пор еще оставался недостаточно выясненным в физиологической литературе. Последнее обстоятельство и побуждает меня представить в настоящей работе анализ субъективных явлений, следующих за разрушением периферических органов равновесия, чтобы таким, образом, насколько позволяют нам данные опытов над животными и наблюдений над человеком, возможно ближе определить специфический характер тех ощущений, которые в нормальном состоянии возбуждаются при посредстве упомянутых органов.
Прежде всего я нахожу, однако, необходимым упомянуть вкратце о тех общих выводах, к которым привело меня изучение рефлекторной деятельности органов равновесия. В этом отношении мы должны иметь ввиду главным образом следующие основные факты и положения.
1. Для равновесия тела служит несколько специально приспособленных периферических органов, в числе которых мы различаем: 1) черепные органы, к которым относятся: а) полукруглые каналы перепончатого лабиринта и в) область воронкообразной части третьего желудочка и 2)внечерепные органы, к которым мы относим нервные аппараты, заложенные в коже и мы шц а х.
Значение полукружных каналов для равновесия тела в настоящее время представляется в такой степени выясненным на основании экспериментальных и патологических наблюдений,, что здесь нет никакой надобности останавливаться на этом предмете. Равным образом и значение области третьего желудочка
1 «Образование наших представлений о пространстве». См. также протоколы Общества психиатров за тот же год.
т
для равновесия тела я считаю выясненным на основании целого ряда опытов, произведенных мною над животными (преимущественно собаками), и некоторых клинических наблюдений. Что касается периферических кожно-мышечных органов, то существование их доказывается, между прочим, следующими фактами. Известно, что если лягушке снять кожу с лапок, то она утрачивает способность поддерживать равновесие тела. Этот известный опыт я повторил над голубем с тем же результатом. Отсюда ясно, что в коже должны содержаться нервные аппараты, которые при посредстве центростремительных путей, поднимающихся в восходящем направлении (по всей вероятности, к мозжечку, по крайней мере у позвоночных высшего типа), обнаруживают резкое влияние на равновесие тела. Эти-то аппараты мы и называем кожными органами равновесия. Более чем вероятно, что и в мышцах содержатся также нервные аппараты, обнаруживающие влияние на равновесие тела, что очевидно по крайней мере из того влияния, которое обнаруживает на равновесие тела нарушение мышечного чувства. Должно заметить, что присутствие на периферии тела особых кожно-мышечных органов равновесия доказывается, между прочим, и тем, что перерезки и патологические процессы в задних столбах спинного мозга приводят к расстройствам в равновесии тела .^сопровождающимся иногда и нистагмом глаз1. При своих опытах "я убедился также, что повреждением боковых частей верхних отделов спинного мозга, в которых заложен так называемый мозжечковый пучок, можно вызвать у животного характеристичные вынужденные движения вокруг продольной оси тела с своеобразным отклонением и нистагмом глаз2.
Здесь необходимо еще заметить, что под именем кожно-мышечных органов равновесия мы понимаем заложенные в коже, в мышцах и, вероятно, в связках периферические нервные аппараты, которые могут быть даже совершенно обособленными от тех чувствительных нервных аппаратов или органов, при посредстве которых воспринимаются осязательные раздражения. На возможность существования на периферии обособленных нервных аппаратов, служащих для функции равновесия тела, я уже указывал в своей работе: «О связи так называемых периферических органов равновесия с мозжечком». В пользу этой обособленности, по моему мнению, говорят с некоторой вероятностью два следующих соображения: 1) закон специфических энергий весьма дурно мирится с тем предположением, чтобы один и тот же нервный аппарат в коже мог служить одновременно и как орган, предназначенный для специальных, т. е. осязательных, ощущений, и как орган равновесия; подобное предположение для
1 См. мою работу «О явлениях, обнаруживающихся при перерезке задних столбов спинного мозга» и пр., Вестник психиатрии, 1890; Pfliiger's Arch., 1890.
2 См. мое исследование «Значение сочетаний методов развития и перерождения» и пр., Неврологический вестник, 1895; Neurol. Zbl., 1895.
122
нас представляется тем более маловероятным, что, как будет выяснено, периферические органы равновесия суть также чувствующие органы, доводящие до сознания определенные ощущения; 2) аналогия с другими периферическими органами равновесия, представляющими собой, как мы знаем, совершенно обособленные от соответствующих органов чувств аппараты, была бы неполною, если принять, что одни и те же органы кожи предназначены и для ощущений прикосновения, и для функции равновесия.
Впрочем, как бы ни был решен вопрос о существовании в коже и мышцах нервных аппаратов для целей равновесия, мы должны иметь в виду, что раздражителями этих органов, по всей вероятности, служат в общем те же самые внешние импульсы, которые являются и раздражителями для специальных органов осязания (прикосновение, давление, растяжение и пр.). Это необходимо принять уже ввиду того влияния, которое оказывает чувство прикосновения и мышечное напряжение на равновесие тела вообще.
2. Все вышеупомянутые «периферические органы равновесия» суть, повидимому, не что иное, как приводы одного и того же центрального механизма, заложенного в мозжечке, в котором импульсы, получаемые от этих органов, передаются на центробежные пути.
Справедливость этого положения, высказанного мною уже в работе «О связи так называемых периферических органов равновесия с мозжечком»1, очевидна как из анатомических отношений периферических органов равновесия к мозжечку, так и из этого обстоятельства, что двигательные явления, обнаруживающиеся при разрушении того или другого из периферических органов равновесия, отличаются замечательным сходством между собой и в свою очередь вполне аналогичны явлениям, следующим за разрушением различных отделов мозжечка.
Этим самым, впрочем, ничуть не исключается возможность и более непосредственной связи упомянутых органов с важнейшими центрами нашего сознания, т. е. с мозговыми полушариями2.
3. Все периферические органы равновесия, служа 'приводами одного и того же центрального механизма, в нормальном состоянии обычно функционируют совместно друг с другом, вызывая импульсы, результатом которых является поддер-
1 Русская медицина, 1884, № 3—5.
2 Анатомические данные о связи периферических органов с мозжечком и последнего с мозговыми полушариями можно найти, между прочим, в моей книге «Проводящие пути мозга», Лейпциг, 1893.
1?Я
жание и исправление нарушенного равновесия.
Это заключение, на которое я указал в своей работе «К физиологии равновесия тела»1, вытекает прямо из того обстоятельства, что возбуждение рефлекторной деятельности всех вообще периферических органов равновесия, если только оно не обусловливается внешними импульсами, передаваемыми при посредстве специальных органов чувств, обычно происходит под влиянием одних и тех же условий, именно при нарушении равновесия тела.
Таким образом, например, при внезапном толчке вправо и нарушении равновесия в этом направлении, вследствие наклона всего тела в правую сторону, приводящего к перенесению центра его тяжести на правую ногу, и обусловленного этим большого сдавливания суставных поверхностей и подошвы правой ноги, приходят в возбуждение, с одной стороны, кожно-мышеч-ные органы правой стороны, с другой —вследствие одновременно происшедшего усиления давления со стороны эндолимфы в левых полукружных каналах возбуждаются также и последние, а вместе с тем приходят в возбуждение, по всей вероятности, и органы, заложенные в правой стенке третьего желудочка (вследствие усиленного давления со стороны черепно-спинной жидкости?). Результатом этих одновременных возбуждений со стороны трех вышеуказанных органов происходит рефлекторная передача импульсов через мозжечок на органы движения, результатом чего и является то непроизвольное напряжение мышц, которое служит к выравниванию нарушенного равновесия тела.
4. Центральный орган равновесия, или мозжечок, находится под непрерывным влиянием импульсов, возникающих в периферических органах равновесия. В мозжечке эти импульсы в свою очередь передаются на двигательные или центробежные приводы, отходящие к мышцам, вызывая, таким образом, отраженные сокращения мышц, направленные к цел и поддержания равновесия тела.
Это общее положение должно быть принято ввиду следующих обстоятельств: обычное положение нашего тела (при движении, стоянии и сидении) в физическом смысле слова представляется в высшей степени неустойчивым, и, не будь центрального, саморегулирующего механизма для равновесия тела, мы, без сомнения, теряли бы его ежеминутно. Последнее, действительно, и происходит вслед за разрушением у животного центрального
1 Военно-медицинский журнал за июнь 1883; Pfluger's Arch., 1883, Bd. XXXI.
124
•органа равновесия —мозжечка, или даже одной пары его периферических приводов (например, полукружных каналов или стенок третьего желудочка). Если, с другой стороны, мы знаем, что всякое нарушение равновесия тела неизбежным образом связано с возникновением импульсов в периферических органах равновесия, то мы и принуждены принять, что при нормальных условиях органы равновесия находятся в непрерывно-деятельном состоянии.
Так как раздражителем одних из этих органов (полукружных каналов и, по всей вероятности, также области третьего желудочка), как мы принуждены пока допустить, служит давление со стороны жидкости на нервные окончания, раздражителем же других (кожно-мышечных органов) являются периферические раздражения, возникающие при всех обычных условиях положения нашего тела (может быть, также в виде давления на нервные окончания в коже и мышцах), то нетрудно понять, что все вообще органы равновесия уже по самому устройству своему представляются в высшей степени приспособленными для возбуждения непрерывного ряда импульсов.
5. Двигательные расстройства, наступающие вслед за разрушением того или другого из органов равновесия, суть прямые последствия дисгармонии в импульсах, получаемых мозжечком от всех вообще периферических органов той и другой стороны.
В работе под заглавием «Результаты перерезки слухового нерва»1 я впервые указал на то обстоятельство, что явления, наступающие при разрушении полукружных каналов одной стороны или при перерезке одного из слуховых нервов, стоят в прямой зависимости как от прекращения функции разрушенных (или отделенных от своего центра) каналов, так и от нормального, непрерывно действующего возбуждения со стороны каналов неоперированной стороны. В другой работе2 я указал, что подобное же объяснение применимо и к явлениям, следующим за разрушением других органов равновесия. Ввиду того, однако, что, как было уже выяснено выше, все периферические органы равновесия суть, повидимому, только приводы одного центрального органа —мозжечка, причем каждый из них составляет только одну часть общего механизма равновесия, и так как, с другой стороны, все эти органы одновременно участвуют в поддержании равновесия тела, вышеуказанное положение относительно явлений, следующих за разрушением того или другого
1 Медицинская библиотека за декабрь 1872; Pfluger's Arch., 1882, Bd. XXX.
1 К физиологии равновесия тела, Военно-медицинский журнал за июнь 1883. Pfluger's Arch., 1883, Bd. XXXI.
125
из органов равновесия, представляется более соответствующим действител ьности.
Согласно с этим положением, непосредственная причина явлений, следующих за разрушением того или другого из органов равновесия, заключается в том, что вместе с уничтожением известного рода импульсов со стороны разрушенного органа нарушается гармония в деятельности всех вообще органов равновесия. Результатом происходящего при этом несоответствия в силе импульсов, возникающих в органах равновесия оперированной и неоперированной стороны, и являются рефлекторные движения подобно тому, как при внезапном нарушении равновесия у совершенно здоровых животных несоответствие в импульсах от органов равновесия той и другой стороны приводит к рефлекторным движениям, служащим к исправлению нарушенного равновесия.
6. Постепенное ослабление двигательных расстройств, обнаруживающихся вслед за разрушением того или другого-из органов равновесия, очевидно, н аходит-ся в связи главным образом с тем, что оставшиеся органы равновесия мало-помалу приспособляются к новым условиям в организме, благодаря чему и устанавливав т-ся относительная гармония в силе импульсов от органов равновесия той и другой стороны.
Справедливость этого положения вытекает из всех наблюдений над животными с разрушением тех или других органов равновесия. Правда, очень распространено мнение, что ослабление двигательных расстройств у животных, которым были разрушены полукружные каналы, обусловливается тем, что раздражение в последних, вызванное операцией, постепенно ослабевает и, наконец, прекращается совершенно; но это объяснение является прямым отголоском теории, объясняющей все вообще симптомы у животных, вызванные перерезкой полукружных каналов, состоянием раздражения в перерезанных каналах. Мне кажется, однако, что в статье «Об отправлении полукружных каналов перепончатого лабиринта»1 я выставил достаточно веские доводы против этой теории и вместе с тем доказал, что явления, наблюдаемые у оперированных животных, обусловливаются главным образом происшедшей вследствие устранения полукружных каналов дисгармонией между нормально притекающими импульсами от органов здоровой стороны и ослабленными импульсами от органов оперированной стороны. Я не имею возможности вновь возвращаться здесь к этому вопросу; замечу лишь, что с теорией возбуждения не вяжется ни необычайная длитель-
1 Международная библиотека, 1882; PHOger's Arch., 1882.
126
ность многих явлений, наблюдаемых у животных (некоторые из них, как известно, остаются даже навсегда, как постоянное явление), ни крайняя медленность и постепенность в улучшении двигательных расстройств, происшедших в результате операции. Единственную опору теория возбуждения находит, собственно, в припадочном течении некоторых из двигательных расстройств, обусловленных разрушением полукружных каналов, как, например, нистагм глаз и вынужденные движения, но припадочный характер этих явлений еще лучше объясняется выставленной мною теорией, имеющей в виду главным образом дисгармонию в импульсах от органов здоровой и оперированной стороны, благодаря чему двигательные расстройства, наблюдаемые у оперированных животных, с одной стороны, являются результатом недостаточного возникновения импульсов в органах оперированной стороны, с другой —результатом перевешивающих по силе эти импульсы возбуждений, исходящих из органов здоровой стороны. Выравнивание этой дисгармонии, очевидно, и происходит путем постепенного приспособления неразрушенных органов равновесия, следовательно: 1)на счет заменяющей функции со стороны оставшейся в целости части оперированного органа равновесия, если таковая имеется, и 2) на счет заменяющей функции со стороны других одновременно функционирующих с ним органов равновесия, благодаря чему происходит медленное и постепенное восстановление нарушенных функций движения и равновесия у животных х.
7. Хотя все периферические орга н'ы р"а в-новесия одинаково служат для целей равновесия, тем не менее они не могут считаться органами равнозначными или тождественными в физиологическом смысле.
Уже в прежних своих работах я указывал на существование связи между каждым из периферических органов равновесия и соответствующим органом чувств. Благодаря этой связи, дана возможность для возбуждения органов равновесия внешними импульсами, передаваемыми при посредстве соответствующих органов чувств —слуха, зрения, кожной поверхности тела и, по всей вероятности, также мышечных сокращений. В этом обстоятельстве, повидимому, и заключается существенное различие относительно рефлекторной функции тех или других периферических органов равновесия. Естественно поэтому, что, хотя двигательные расстройства, следующие за разрушением одного из этих органов, и могут со временем в более или менее значительной
1 Вместе с этим, без сомнения, не исключается и возможность влияния раздражения со стороны операционной раны на движения животного; но мы признаем во всяком случае, что это влияние далеко не играет той видной роли, которое ему обыкновенно приписывают, и если оно вообще и обнаруживается при операциях над органами равновесия, то лишь в первые дни после произведенного разрушения.
127
степени ослабеть под влиянием заменяющей функции других подобных же органов, но рефлекторное влияние внешних импульсов с соответствующего органа чувств на механизм равновесия при посредстве разрушенного периферического органа должно быть уже утраченным навсегда.
Ознакомившись со всеми приведенными данными, относящимися до рефлекторной функции органов равновесия, нам будет легче уяснить себе значение этих органов в другом отношении.
В начале работы я упомянул, что предпринятые мною исследования относительно периферических органов равновесия привели меня к заключению, что эти органы, помимо своей рефлекторной функции, служат еще источником определенных ощущений. Справедливость этого вывода может быть доказана на животных следующим образом:
Представим себе, что у данного животного мы произвели обширное разрушение одного из органов равновесия, например, всех полукружных каналов, или слухового нерва, или повреждение задне-боковой части стенки третьего желудочка на одной стороне. Вслед за этой операцией животное обычно начинает производить вынужденные вращательные движения вокруг продольной оси тела, причем со стороны глазных яблок обнаруживается своеобразное скошение (одно —обращено кверху и кнаружи, другое — книзу и кнутри), сопровождаемое резким нистагмом. По истечении некоторого времени вращение прекращается и животное принимает покойное положение на той стороне туловища, в направлении которой происходило вращение. При этом замечаются нистагм и упомянутое отклонение глазных яблок, голова и передняя часть туловища поворочены вокруг продольной оси в направлении бывшего вращения, противоположные конечности вытянуты и напряжены, соответствующие же подогнуты и легко поддаются пассивным сгибаниям и разгибаниям. Спустя короткое время (обыкновенно не более нескольких минут) вращение снова развивается с прежнею силою, возникая или самопроизвольно, или под влиянием известных внешних раздражений (толчка, испуга, болевого раздражения и пр.).
Такова последовательность явлений в первые дни вслед за операцией; дальнейший же ход двигательных расстройств и постепенное их выравнивание здесь могут быть оставлены без внимания.
В своих прежних работах я настойчиво указывал, что все описанные явления суть по преимуществу рефлекторной природы. Действительно, та же самая операция у животных, которым предварительно были удалены мозговые полушария, обыкновенно вызывает подобные же двигательные расстройства, как и у животных с неповрежденными полушариями. Было бы, впрочем, ошибочно думать, что операция удаления мозговых полушарий вовсе не обнаруживает влияния на двигательные расстройства, являющиеся последствием разрушения органов равновесия.
J 28
Напротив того, если у животного с вращением вокруг оси тела мы удалим оба мозговые полушария, то вслед за этой операцией нистагм глаз и приступы вращения всегда резко ослабевают и часто уже не появляются иначе, как под влиянием тех или других внешних раздражений; своеобразное же отклонение глазных яблок, поворачивание головы и передней части туловища вокруг продольной оси тела, а равно и вынужденное положение на одной стороне туловища при этом еще обнаруживаются почти с прежнею силою.
Указанное ослабление двигательных расстройств в данном случае, без сомнения, легче всего объяснить тем, что вместе с удалением мозговых полушарий устраняется ряд импульсов, выходящих из этого органа и побуждающих животное к продолжению вращательных движений. Это заключение, в свою очередь, заставляет нас принять, что разрушение органов равновесия не только вызывает у животных рефлекторные двигательные расстройства, но и действует определенным образом на психическую сферу, вы з ы в а я в ней изменения, которые до известной степени служат моментом, усиливающим рефлекторные двигательные расстройства.
Спрашивается, какие же изменения психической сферы могут быть результатом разрушения органов равновесия?
Ответ на этот вопрос дают нам наблюдения поражений мозжечка или тех или других его приводов, resp. периферических органов равновесия, опыты с электризацией в затылочной области головы у человека и, наконец, опыты с пассивным и активным вращением вокруг оси тела.
Уже с давних пор известно, что поражение лабиринта, мозжечка и его ножек у человека, кроме расстройств в равновесии тела, вынужденных движений и явлений нистагма глаз, сопровождается в высшей степени резкими явлениями головокружения. Известны также случаи поражения продолговатого мозга в области нижних олив, связанных теснейшим образом с мозжечком и, повидимому, также с областью третьего желудочка при посредстве описанного мною центрального пучка покрышки; в этих случаях рядом с характеристичными двигательными расстройствами обнаруживались также приступы резкого головокружения. Наконец, в работе «Об отправлении центрального серого вещества третьего мозгового желудочка» я привел случаи поражения названной области, в которых также с постоянством замечались явления сильного головокружения.
В настоящее время вообще можно считать доказанным, что головокружение при поражении мозжечка и его приводов составляет симптом столь же характеристичный, как и двигательные расстройства в виде нарушения равновесия тела. Многие из
Э В. М. Бехтерев
129
больных в состоянии с подробностью описать это головокружение и говорят в таком случае или о кажущемся отсутствии прочной опоры под ногами, или о ненормальном ощущении движения собственного тела в известном направлении: то в виде падения вперед или назад, то в виде вращения в ту или другую сторону. К этим явлениям нередко присоединяется еще кажущееся движение всех окружающих предметов, которое, по моим наблюдениям, обычно происходит в сторону, противоположную в отношении направления субъективного движения тела.
Явления, обнаруживающиеся при гальванизации в затылочной области головы у человека, были уже известны Пуркинье 1 и с тех пор изучались различными наблюдателями; в особенности же Бреннеру2 и Гитцигу3 мы обязаны наиболее обстоятельными работами в этом направлении. Не имея возможности входить здесь в подробное обсуждение всех сюда относящихся исследований, я ограничусь только сжатым описанием самих явлений, имея при этом в виду наблюдения, сделанные над самим собою.
Если электроды гальванического тока приставлены к затылочной области головы, вблизи сосцевидных отростков, то в момент замыкания цепи и во все время прохождения тока мы испытываем ряд ощущений в виде сильного головокружения. Последнее состоит в ложном ощущении движения собственного тела в направлении положительного полюса или анода и в кажущемся движении всех Еообще видимых предметов в противоположном направлении. Если мы в эту минуту закроем глаза и прикоснемся к какому-либо из окружающих нас предметов, то осязаемый предмет нам кажется движущимся в сторону катода. Наконец, если во время опыта в комнате помещается какой-либо предмет, издающий однообразный и продолжительный шум (например, дребезжащий стук молоточка от индукционного аппарата, шум газовой горелки и пр.), то и самый источник шума в случае более сильного головокружения (в особенности при закрытых глазах) кажется как бы удаляющимся от нас в том же направлении, как видимые и осязаемые предметы.
Объективно при этом наблюдается действительное и нередко в высшей степени резкое движение тела в сторону положительного полюса или анода и отклонение глаз с нистагмом в противоположном направлении.
Размыкание цепи сопровождается вообще явлениями тождественного характера, но менее продолжительными и противоположными по направлению.
1 Ruest's Magazin f. d. ges. Heilkunde etc., Berlin, 1827, Bd. XXIII; H i t z i g, Untersuchungen iiber das Gehirn, Berlin, 1874, S. 197.
8 Brenner, Untersuchungen und Beobachtungen auf dem Gebiete der Elektrotherapie, Leipzig, 1868, d. I и II.
3 H i t z i g, Reichert's η. Du Bois-Raymond's Arch., 1871, Hft. 5u. 6; Untersuchungen iiber des Gehirn, Berlin, 1874.
130
Если опыт делается при закрытых глазах, то субъективное ощущение движения тела еще более усиливается, действительное же его движение обнаруживается во всяком случае с неменьшей резкостью, как и при открытых глазах. В обоих случаях ко всем упомянутым явлениям присоединяется еше в высшей степени неприятное чувство тошноты. Связь всех выше описанных явлений с нарушением функции полушарий мозжечка или его приводов вполне очевидна и вообще не может подлежать какому-либо сомнению. Между различными наблюдателями по этому поводу возникали только разногласия совершенно частного характера. Так, например, Бреннер и Мах, вопреки другим авторам, думали свести описанные выше яиления на раздражение-одних полукружных каналов. Впоследствии, однако, это мнение было сильно пошатнуто возражениями Гитцига. Наконец, не очень давно Шпамер х показал, что и после удаления всех перепончатых каналов у голубей еще удается наблюдать вращение при поперечном прохождении тока через голову. Новейшие исследования с электризацией мозжечка, произведенные над глухими, у которых явлений головокружения при этом будто бы не обнаруживается, снова говорят в пользу того, что в происхождении этих явлений, действительно, участвуют полукружные каналы; тем не менее, ввиду вышеприведенных опытов Шпамера, вряд ли можно сомневаться в том. что и мозжечок играет известную роль в происхождении этих явлений. Для нашей цели, впрочем, безразлично, будет ли окончательное решение этого вопроса в том или ином смысле, так как мы знаем, что волокна полукружных каналов проходят в мозжечок и сами каналы представляют собою не что иное, как один из периферических приводов мозжечка. Мы заметим лишь, что непосредственная причина наблюдаемых расстройств при гальванизации затылочной области головы, без сомнения, заключается в различном влиянии катэлектротона и анэлектротона на нервные приводы. Неопровержимым доказательством этого служит, между прочим, тот факт, что если к сосцевидным отросткам приставить одинаковые полюсы тока (для чего удобнее гсего пользоваться раздвоенным проводником), противоположный же полюс отвести к безразличной части тела, например, к задней части шеи или к грудине, то уже не удается наблюдать вышеописанных явлений (Бреннер, Гитциг).
Явления, обнаруживающиеся при вращении вокруг оси тела, как увидим ниже, также находятся в непосредственной связи с нарушением функции органов равновесия. Так как эти явления хорошо известны всякому наблюдателю, то здесь нам достаточно будет упомянуть о них только вкратце. !
Если мы сделаем несколько быстрых Еращательных движений вокруг оси тела, то в момент остановки мы почувствуем, как буд-
1 Spamer, Pfluger's Arch., 1878, Bd. XXI, Hft. 10—12.
9*
131
то?бы тело каше внезапно получило сильный толчок в противоположном направлении. Вместе с тем все видимые нами предметы кажутся движущимися в сторону бывшего вращения. Закрывши глаза, мы можем убедиться, что подобный же обман замечается и в органах осязания и слуха. Так, если мы прикасаемся к какому-либо из окружающих предметов, то мы чувствуем, что он движется под нашими пальцами в сторону бывшего вращения; точно так же источник шума, если последний продолжается известное время, кажется нам удаляющимся в том же направлении.
Вместе с ощущением толчка или кажущегося падения в направлении, противоположном вращению, мы, действительно, непроизвольно отшатываемся в этом направлении и можем даже упасть, если не успеем во-время предупредить это падение соответствующими телодвижениями. Глаза при этом отклоняются в сторону вращения и обнаруживают нистагм. Совершенно подобные же явления замечаются и в том случае, если вращение производят с закрытыми глазами и открывают их только в момент остановки; если же глаза остаются закрытыми и после вращения, то чувство падения в противоположном направлении для нас становится еще более ясным, и при этом мы почти неизбежно падаем, если только не успеем во-время ухватиться за какой-либо предмет.
После более или менее продолжительного вращения вместе с сильным головокружением почти всегда развивается в высшей •степени тягостное чувство тошноты и в то же время видимые предметы нам кажутся неясными, как бы со стертыми краями. Дело может дойти, наконец, до потемнения зрительного поля в такой степени, что остается, повидимому, только субъективное ощущение света, при этом слух и осязание также притупляются весьма заметным образом. Слабые звуки в этом состоянии почти вовсе не достигают до нашего сознания, более сильные — субъективируются, а шероховатая поверхность нам кажется гладкою.
Как эти последние явления, так и все вообще симптомы головокружения в только что указанном случае довольно быстро исчезают под влиянием нескольких оборотов в направлении, противоположном прежнему вращению тела и, следовательно, в направлении, противоположном кажущемуся движению окружающих предметов.
Со своей стороны я замечу, что и в случаях головокружения, вызванного гальванизацией затылочной области головы, а равно и при головокружении, вызванном патологическими процессами, оно всегда заметным образом ослабевает при вращении тела и вообще при движениях, противоположных кажущемуся движению предметов.
Явления, наблюдаемые при пассивном вращении тела, были с подробностью изучены пражским физиком Махом 1. Мы за-
1 М а с h, Crundlinien d. Lehre von den Bewegungsernpfindungen, Leipzig, 1875.
132
имствуем здесь только наиболее интересные для нас результаты его исследования.
Подвергаемый вращению наблюдатель, если он помещен в бумажном ящике, следовательно, вполне изолирован от зрительных впечатлений, при начале вращения обыкновенно еще в состоянии определить его направление. При дальнейшем же вращении это ощущение утрачивается и вместе с тем появляется субъективное чувство вращения в противоположном направлении. Вслед за внезапной приостановкой вращения это субъективное чувство движения или вращения еще более усиливается. Если в этот момент быстро открывают бумажный ящик, то все окружающее нас пространство представляется также вращающимся. Впечатление, получаемое при этом, может быть выражено следующим образом: «кажется, как будто все видимое пространство вращается в другом неподвижном, хотя и видимом» *. При этом ось вращения всегда совпадает с положением нашей головы, что было известно уже по опытам Пуркинье и Дарвина (старшего) 2.
Все только что описанные явления, наблюдаемые при активном и пассивном вращении тела, без сомнения, имеют полнейшую аналогию с явлениями, наступающими при гальванизации в затылочной области головы. В свою очередь, и те, и другие явления поражают своим сходством с явлениями, обнаруживающимися в патологических случаях поражения мозжечка или принадлежащих ему периферических проводников. И там, и здесь наблюдаются вынужденные движения тела с отклонением глаз и явлениями нистагма. Точно так же и характер головокружения в обоих случаях отличается полнейшим сходством.
Таким образом, не может подлежать сомнению, что явления, наступающие при активном и пассивном вращении вокруг оси тела так же, как и явления, наблюдаемые при гальванизации в затылочной области головы, находятся в зависимости от нарушения функции органов равновесия, следовательно, мозжечка, или его периферических приводов.
Некоторые авторы, как Мах и Бреннер, допускавшие зависимость явлений, обнаруживающихся при гальванизации в затылочной области головы, от нарушения функции полукружных каналов, также и явления, наступающие вслед за вращением вокруг оси тела, старались объяснить расстройством функции одних каналов, происходящим под влиянием перемен в давлении содержащейся в них жидкости. С тех пор, однако, как мы знаем о существовании других периферических органов равновесия,
1 М ас h, loc. cit., p. 26.
2 Хотя Max в своем описании не делает указаний относительно объективных явлений (как отклонение тела, изменения в положении глаз и др.), но, без всякого сомнения, эти явления должны были обнаруживаться в его опытах так же, как и при активном вращении вокруг оси тела.
133-
по функции совершенно сходных с полукружными каналами, подобный взгляд, без сомнения, уже не может быть более защищаем. Против него, впрочем, имеются и фактические данные, так как у животных с перерезанными слуховыми нервами при вращении удается еще наблюдать все те объективные явления (отклонение тела и головы, нистагм глаз и пр.), которые обнаруживаются при вращении здоровых животных.
Ввиду этих данных явления, наблюдаемые при вращении вокруг оси тела, мне кажется, правильнее рассматривать как явления сложной природы, находящиеся в зависимости от нарушения функции всех вообще органов равновесия, следовательно, и полукружных каналов, в области третьего желудочка и кожно-мышечных органов. Возможно думать также, что и мозжечок принимает некоторое участие в обнаружении указанных явлений, так как вследствие развития центробежной силы при вращении тела, как предполагал уже Пуркинье, полушария мозжечка могут подвергаться сдавливанию, равным образом и при гальванизации затылочной области головы ток может достигать не только до полукружных каналов, но и мозжечковых полушарий.
Все приведенные выше факты убеждают нас в том, что при самых различных влияниях, действующих па органы равновесия, кроме объективных явлений, обнаруживаются еще в высшей степени резкие изменения психической сферы в виде характеристического головокружения. Естественно поэтому заключить, что у животных при повреждении мозжечка или его приводов рядом с рефлекторным расстройством движения должны существовать подобные же субъективные явления в психической сфере, которые в свою очередь не остаются без влияния на двигательную сферу, усиливая наклонность к продолжению вынужденных движений.
Короче, явления, следующие за разрушением полушарий мозжечка или его периферических приводов как у человека, так и у животных, распадаются на два отдела: 1) объективные явления, состоящие в расстройстве равновесия тела и в непроизвольных или вынужденных движениях, сопровождаемых своеобразным отклонением глаз и явлениями нистагма; 2) субъективные явления в виде головокружения, состоящего в ложном ощущении движения собственного тела в определенном направлении, именно — в направлении действительного непроизвольного или вынужденного движения тела — и вместе с тем кажущемся движением в противополож-
!;м
ную сторону всех окружающих предметов, воспринимаемых нами при посредстве органов зрения, осязания и слуха.
Перейдем теперь к более подробному анализу и взаимной связи этих явлений.
В физиологии, как известно, существовала теория, по которой двигательные расстройства, в особенности же вращение животных с повреждением мозжечка или каналов, объяснялись развитием головокружения, иначе говоря, объективные явления ставились в прямую зависимость от явлений чисто субъективного свойства. При этом одни авторы, как Грасиоле и Левая *, допускали, что развитие головокружения у оперированных животных стоит в связи с своеобразным отклонением глаз; по мнению же других, это головокружение имеет своим источником полушария головного мозга 2.
Подобная теория, однако, не может быть нами принята главным образом ввиду следующих фактов:
1. Я убедился, что у оперированных животных с вращением вокруг продольной оси последнее не прекращается и вслед за вылущением глазных яблок 3.
2. Удаление мозговых полушарий у подобных животных, как мы упоминали, вызывает только известное ослабление двигательных расстройств; все же характеристические явления у них еще обнаруживаются и вслед за этой операцией.
3. Наконец, мы знаем, что в патологии мозжечка существуют такие случаи, в которых обнаруживаются вполне ясные двигательные расстройства при совершенном отсутствии головокружения.
Нельзя согласиться также с мнением Гитцига 4 и Вундта5, по которым вынужденные движения, обнаруживающиеся при гальванизации затылочной области головы, должны быть рассматриваемы как движения произвольные, направленные к исправлению кажущегося нарушения равновесия.
При всех моих опытах с гальванизацией в затылочной области головы так же, как и в опытах с вращением вокруг оси, отклонение тела всегда происходило в направлении субъективного движения и, следовательно, никак не могло служить к цели исправления равновесия. Кроме того, указанное отклонение тела мы всегда сознаем уже как совершившийся факт, не имея никакой возможности предупредить его или задержать. Последнее, очевидно, также мало говорит в пользу того, чтобы вынуж-
1 Gratiolet et Leven, Mouvements de rotation sur l'axe determines par les lesions du cervelet, L'Institut, I860, v. 28.
2 V u 1 p i a η, Lecons sur la physiologie gen. et сотр., Paris, 1866, p. 599 et suiv.
3 На этот факт указывает также Вульпиан (там же, стр. 596). « Hitzig, Unters. uber das Qehirn., S. 225.
* Вундт В., Основы физиологической психологии, СПБ, вып. 1.
135
денные движения зависели от нашей воли. Наконец, больные с поражением мозжечка или полукружных каналов никогда не определяют вынужденные движения как произвольные. Многие из них вовсе не могут дать ясного отчета о происхождении этих движений, тогда как другие для обозначения их употребляют стереотипное выражение: «как будто бы какая-то невидимая сила постоянно их толкала в известном направлении»".
Приводя эти данные, я вовсе не имею в виду совершенно отрицать влияние произвольных импульсов на движения, обнаруживающиеся при указанных условиях. Я хотел только сказать, что упомянутые движения, так же как и движения животных с разрушением органов равновесия, носят по преимуществу рефлекторный характер, но, без сомнения, кажущееся движение видимых предметов, 'а равно и субъективно сознаваемое непроизвольное движение тела, обманывая относительно положения собственного тела в окружающем мире, в свою очередь может служить причиной произвольного импульса, способствующего движению тела в том же направлении, в котором совершается и непроизвольное или рефлекторное вынужденное движение.
Следовательно, произвольные движения в данном случае являются движениями вторично обусловленными, тогда как непосредственно вызванные вышеуказанными влияниями движения всегда носят на себе характер движений непроизвольных или рефлекторных, вследствие чего они и понимаются как движения вынужденные или насильственные.
Но если двигательные расстройства, обнаруживающиеся при нарушении функции органов равновесия, как мы только что выяснили, носят по преимуществу рефлекторный характер и только отчасти могут зависеть от субъективных явлений головокружения, то, быть может, не следует ли рассматривать эти последние как результат появляющихся у животного двигательных расстройств.
Действительно, некоторые из авторов высказывались в том смысле, что головокружение, появляющееся при нарушении функции органов равновесия, находится в прямой зависимости от движения глазных яблок. Вот, например, как Гитциг в своем классическом труде говорит о причине головокружения при гальванизации мозжечка: «Наступление такого рода кажущегося движения при описанных движениях глазных яблок представляет в такой мере физиологический постулат, что объяснение этой части головокружения движениями глаз я считаю не только вероятным, но и необходимым».
На мой взгляд, однако, все подобные теории не могут считаться вполне удовлетворительными. Дело в том, что хотя и могут быть приведены неоспоримые доказательства в пользу возможности развития головокружения под^ влиянием непроиз-
136
больных движений глазных яблок1, но не в этом, повидимомуг заключается главная причина головокружения, наступающего-как при вращении тела, так и под влиянием гальванизации мозжечка, а равно и вообще при нарушении функции органов равновесия.
Мы знаем, в самом деле, что больные, страдающие периферическим нистагмом, уже вскоре после развития своей болезни не испытывают головокружения. Далее известно, что при головокружении, развивающемся под влиянием вращения вокруг оси тела, фиксирование глаз не прекращает кажущегося движения окружающих предметов. Некоторые авторы говорят даже об усилении головокружения при этом условии (Мах, Цион). Здесь уместно привести также интересный опыт Маха: если при вращении вокруг оси тела давлением на глазное яблоко вызвать в поле зрения фосфен, то оказывается, что последний участвует в кажущемся движении предметов даже и при том условии, если глаз фиксирован прочно и вследствие того все время остается неподвижным.
Бреннер, впрочем, и этот факт старался объяснить с точки зрения вышеупомянутой теории, допуская, что фосфен смещается в таких случаях под влиянием только одних иннерва-ционных ощущений, хотя бы при этом и не происходило действительного движения глазного яблока. Но Цион2 мог наблюдать смещение фосфена даже и в том случае, если во время опыта при закрытых глазах фиксируют близко отстоящий палец или кончик носа. Без сомнения, при подобном условии ощущения иннервации, как замечает сам Цион, могут дать только представление о неподвижности глаз и, следовательно, зрительное головокружение может происходить и без активного участия зрительных органов.
Здесь нельзя упустить из виду еще следующего обстоятельства: как известно, при вращении вокруг оси тела наступает не одно только зрительное головокружение, но также осязательное и слуховое; вряд ли при этом можно сомневаться в том, что все эти ложные ощущения, происходящие в различных органах
1 Примером могут служить, например, случаи развития головокружения, появляющегося при смотрении на движущиеся перед глазами предметы, на быстрое течение водопада и пр. В работе «К физиологии равновесия тела» (Pfluger's Arch., 1883, Bd. XXXI) я объясняю подобное головокружение непосредственным влиянием движений глазных яблок на функцию области третьего желудочка. Ввиду признания в настоящее время волокон, выходящих из зрительных нервов позади chiasma непосредственно в серое вещество области третьего желудочка (см. мои «Проводящие пути мозга»), можно думать и о возможности передачи при посредстве этих волокон к органу равновесия, заложенному в области третьего желудочка, самих зрительных импульсов, независимо от всяких движений глазных, яблок.
2 Ц и о н, Об отправлении полукружных каналов и т. д., Военно-медицинский журнал за июль 1879; также These pour le doctorat, Paris, 1878.
13?
•чувств, должны быть результатом одной и той же общей причины. Спрашивается теперь, как же мы должны объяснить происхождение осязательного и слухового головокружения при вращении тела, если кажущееся движение окружающих предметов мы поставим в зависимость от неправильного отклонения глазных яблок?
Наконец, я должен указать на многочисленные, известные всем клиницистам, факты из патологии человека, доказывающие, что при поражениях мозжечка и полукружных каналов характеристическое головокружение нередко обнаруживается при отсутствии каких-либо объективных явлений. Точно так же в приведенных выше опытах с вращением вокруг оси тела и в опытах с электризацией в области мозжечка, если только применяют слабый ток, явления головокружения обнаруживаются прежде, чем удается заметить какие-либо изменения в положении глазных яблок.
Все эти данные приводят нас к тому выводу, что симптом головокружения в указанных случаях в такой же мере составляет прямое последствие нарушения функции органов равновесия, как и своеобразные двигательные расстройства. Последние, быть может, служат только моментом, усиливающим до известной степени приступы головокружения, но ничуть не составляют главной причины его развития, как допускали некоторые авторы.
Итак, объективные и субъективные явления, обнаруживающиеся при нарушении функции или при разрушении органов равновесия, не относятся друг к другу как причина к следствию. Оба ряда явлений представляют собою только сочетанный симптом, будучи в то же время прямым следствием одной и той же причины. При разрушении органов равновесия, следовательно, мы имеем явления, совершенно аналогичные тому, что наблюдается при поражении других чувствующих органов: с одной стороны, прекращение известного ряда рефлексов, источником которых служит данный орган, с другой — нарушение восприятия определенных ощущений. Это в свою очередь говорит в пользу того, что органы равновесия не только служат в организме постоянным источником рефлекторных движений, направленных к цели поддержания равновесия тела, но •они должны быть также рассматриваемы и как органы, воспринимающие известные ощущения.
Здесь мы займемся еще на короткое время характером субъективных явлений, обнаруживающихся при тех или других влияниях на органы равновесия, а затем перейдем к выяснению тех •ощущений, которые воспринимаются этими органами.
Головокружение, обнаруживающееся при нарушении функции органов равновесия, как мы видели, состоит, собственно
J 38
говоря, из двоякого рода ложных ощущений: 1) из ощущений движения собственного тела в известном направлении и 2) из кажущегося движения в противоположном направлении всех предметов, которые мы в состоянии видеть, осязать и звуки которых мы слышим.
Спрашивается теперь: в каком отношении состоят друг к другу оба ряда ощущений и не существует ли между ними внутренней связи?
Я думаю, что в этом случае мы, действительно, имеем тесную связь одного явления с другим, т. е. что одно из них находится в прямой зависимости от другого. Это ясно уже из того обстоятельства, что ощущение движения собственного тела и кажущееся движение окружающих предметов прямо противоположны по направлению.
Известно, что пассивные движения своего тела мы обыкновенно переносим на окружающий нас мир, причем все предметы нам кажутся движущимися в противоположном направлении. •С другой стороны, и движение окружающих предметов мы нередко переносим на самих себя. Достаточно припомнить здесь всем известный пример, что во время остановки на станции железной дороги, при передвижении поездов мы нередко поражаемся следующим явлением: нам кажется, что видимые нами поезда стоят на одном месте, тогда как наш вагон пришел в движение.
Таким образом, легко представить себе, что если при известных влияниях на органы равновесия мы получим с самого начала ложное ощущение движения собственного тела в известном направлении, то вместе с тем все окружающие предметы нам должны казаться движущимися в обратном направлении. Наоборот, если бы при тех же условиях мы получили первичное ощущение в виде движения окружающих предметов, то мы перенесли бы его на самих себя и таким образом ощущали бы вместе с тем движение собственного тела.
Для нас важно поэтому выяснить — какое из двух упомянутых ощущений в данном случае мы должны считать первичным или основным? Этот вопрос представляется возможным разрешить на основании следующего соображения.
Если бы кажущееся движение окружающих предметов при нарушении функции органов равновесия было ощущением первичным или основным, то, без сомнения, как при опытах с вращением вокруг оси тела, так и при гальванизации в затылочной области головы ложное ощущение движения собственного тела должно было бы исчезать или по крайней мере уменьшаться в более или менее резкой степени вместе с закрытием глаз. Между тем опыт, как мы видели, показывает, что при подобных условиях чувство движения собственного тела не только не уменьшается, а, наоборот, еще в значительной степени усиливается.
139
Последнее обстоятельство заставляет нас с положительностью заключить, что при влияниях, действующих на органы равновесия, основным ощущением является чувство движения собственного тела в известном направлении, кажущееся же движение окружающих предметов в противоположную сторону есть неболее, как ощуще ниевтор и чное, составляющее прямое последствие первого ощущения.
Итак, различные нарушения в деятельности органов равновесия вместе с рефлекторными двигательными расстройствами возбуждают прежде всего ощущения движения тела в том или другом направлении. Выяснив этот факт, нам уже не трудно будет определить и специфическую энергию рассматриваемых органов, а вместе с тем выяснить себе и характер тех ощущений, которые в нормальном состоянии воспринимаются при посредстве этих органов.
Некоторыми авторами уже были предприняты попытки определить специфический характер ощущений одного из органов равновесия, именно —полукружных каналов. Упомянем здесь прежде всего о двух более распространенных мнениях Гольца, Маха и Бреннера. Первый автор высказался в том смысле, что полукружные каналы представляют собою чувствительные органы для равновесия головы и косвенным образом всего тела («sind, so zu sagen, Sinnesorgane fur das Gleichgewicht des Kop-fes tmd mitlelbar des ganzen Korpers»).
Нетрудно видеть, что подобное определение имеет в виду явления нарушения равновесия, следующие за разрушением каналов. Мы знаем, однако, что эти явления суть по преимуществу рефлекторной природы и не стоят в прямой зависимости от каких-либо ощущений.
Спрашивается поэтому, существует ли какая-либо возможность определять по этим явлениям специфический характер тех ощущений, которые воспринимаются при посредстве органов равновесия? Конечно, нет. В пояснение сказанного представим себе следующий пример: вместе с перерезкой зрительного нерва,. как известно, вполне прекращается рефлекторное сокращение зрачка под влиянием света; но можно ли заключать отсюда, что зрительный нерв представляет собою чувствующий орган для поддержания радужной оболочки в сокращенном состоянии?
Гипотеза Маха и Бреннера, без сомнения, представляет уже значительный шаг вперед, хотя и она имеет свои слабые стороны.
По мнению Маха, полукружные каналы суть органы, при посредстве которых мы определяем движение собственного тела и, следовательно, специфические ощущения этих органов суть ощущения движения тела. Главное основание, на которое опирается этот взгляд, очевидно, имеет в виду одно из положений
140
автора, а именно: как бы мы ни изменяли во время пассивных вращений характер тактильных и мышечных ощущений наблюдателя, последний тем не менее еще сохраняет способность определять направление движения. Отсюда автор и заключает, что в организме нашем должен существовать особый специфический орган, при помощи которого мы определяем направление движений нашего тела. Путем дедукции Мах приходит к заключению, что этот орган должен представлять собою полукружные каналы. В более полном развитии теории Маха и Бреннера, приписывая полукружным каналам задачу воспринимать ощущения вращения и движения тела вообще, признает вместе с тем за отолитами функцию, благодаря которой мы ориентируемся в пространстве. То и другое выполняется благодаря следующим условиям: каждый полукружный канал, как известно, представляет собою дугообразную полость, наполненную эндолим-фой. При всяком вращении головы, благодаря инерции, происходит обратное относительное движение жидкости, передающееся на выступающие в просвет нервные волоски, служащие нервными окончаниями п. vestibuli; благодаря этому механическому раздражению нервных окончаний, и сознается вращение жидкости, а вместе с тем и вращение головы. Так как далее человек обладает тремя полукружными каналами, расположенными в направлении трех перпендикулярно пересекающихся плоскостей, то, очевидно, таким образом при посредстве полукружных каналов воспринимается вращение вокруг каждой оси тела. С другой стороны, нервные окончания отолитов, как известно, вступают в желатинозную массу, образуя с последней пластинку, на которой покоится отолит. Каждое ускорение движения вызывает смещение отолита как более тяжелого тела, причем смещение отолита также производит механическое влияние на подлежащие нервные волоски. Таким образом, движения отолитов, механически раздражая нервные окончания, в свою очередь, дают нам возможность заключать о движении головы. Должно заметить, что теория эта подкрепляется опытными исследованиями над глухонемыми со стороны Джэмса и Крейдля1, но лишь в отношении влияния полукружных каналов на развитие головокружения и рефлекторных движений глаз, появляющихся при вращении тела вокруг оси. Это влияние, без сомнения, должно признать бесспорным, хотя уже из вышеприведенных данных представляется очевидным, что, кроме полукружных каналов, на развитие тех и других явлений обнаруживают влияние и деятельность других органов равновесия, по функции совершенно аналогичных полукружным каналам. Вышеприведенную гипотезу Маха и Бреннера мы считаем ошибоч-
1 S. А. К г е i d 1, Zur physiol. Bedeutung d. Ohrlabyrinthes, Wiener klin. Wschr., 1892, №2; ProtoK. d. K- K- Gescllsch. d. Aerzte in Wirnam, 12 Febr. 1892.
141
ной в той части, которая касается объяснения субъективных, явлений, обусловливаемых функцией полукружных каналов. Дело в том, что представление о движении нашего тела мы можем составить себе не иначе, как при посредстве ощущений положения его в пространстве. Без подобных ощущений немыслимо было бы и представление о движении тела. Иначе говоря, прежде чем ощущать движение тела, мы должны ощущать его положение.
Что ощущения положения тела действительно присущи нашему организму, мне кажется, не может подлежать сомнению. Известно, что в каких бы условиях равновесия мы ни находились, мы всегда в состоянии определить с известной точностью положение своего тела даже в том случае, если при этом мы не пользуемся ни зрительными, пи осязательными или мышечными ощущениями. Так, при пассивном погружении в воду с закрытыми глазами мы все еще в состоянии с известной точностью определить в каждую данную минуту как направление, так и величину уклонения своего тела по отношению к вертикальной линии. Вряд ли, кроме того, можно сомневаться в том, что в подобных случаях мы ощущаем прежде всего положение своей головы;, определение же положения других частей тела в отношении головы, очевидно, может быть уже достигнуто при посредстве ощущений, исходящих с нашей кожной поверхности и от мышц.
В действительности органы равновесия по своему устройству представляются в высшей степени хорошо приспособленными для ощущений положения головы, а вместе с тем и всего тела.
Если принять, что давление жидкости в полукружных каналах и в воронкообразной части третьего мозгового желудочка служит естественным раздражителем для нервных окончаний этих органов, в пользу чего, по моему мнению, существуют вполне веские соображения, то легко представить себе, что, сообразно положениям нашей головы, давление на те или другие чувствительные приборы, заложенные в стенках упомянутых органов, должно изменяться, что и служит нам источником специфических ощущений положения головы.
Что касается кожно-мышечпых органов равновесия, нормальным раздражителем которых, как следует думать, является прикосновение и вообще давление на кожную поверхность и суставы и растяжение мышц и связочного аппарата, то, очевидно, они более приспособлены к определению положения отдельных членов по отношению к положению головы. Весьма вероятно, что при посредстве этих именно органов мы и воспринимаем специфические ощущения положения своих членов, которые мы относим к так называемому мышечному чувству. Во всяком случае, ощущения, возникающие при посредстве кожно-мышечных органов равновесия, должны служить прямым дополнением к тем ощущениям поло-
142
жения головы, которые мы получаем на основании импульсов от полукружных каналов и области третьего желудочка.
Само собою разумеется, что эти ощущения положения головы и прочих частей тела, воспринимаемые при посредстве органов равновесия, лежат, между прочим, и в основе того чувства, которое мы называем чувством равновесия тела. С этой стороны Гольц, признававший в полукружных каналах чувствительные органы для равновесия головы и косвенным образом всего тела, без сомнения, был весьма близок к истине.
Допустив, что периферические органы равновесия служат нам прежде всего источником особых ощущений положения головы и тела вообще, мы вместе с тем должны'принять, что при посредстве тех же самых органов мы определяем и перемену этого положения, т. е. движение тела. Дело в том, что при всяком движении тела в наших органах равновесия должен возникать последовательный ряд импульсов, на основании которых создаются ощущения положения тела в различные промежутки времени. Это и дает нам возможность иметь представление о движении нашего тела.
Совершенно подобно тому, как наша сетчатка, будучи приспособлена для восприятия зрительных впечатлений, в состоянии определять не только положение окружающих предметов, но и движение их, так точно и наши органы равновесия, приспособленные для восприятия ощущений положения головы и прочих частей тела, в состоянии определять также и движение последнего, а равно и скорость этого движения.
Необходимо, впрочем, иметь в виду, что наша способность определять скорость движения тела представляется, вообще говоря, довольно ограниченною. Мах доказал своими опытами, что мы в действительности не способны определять постоянную скорость движения тела, а замечаем только ускорение этого движения. Следовательно, только колебания в скорости движения тела могут служить источником ощущений, по которым мы определяем направление этого движения1. Крайне медленное ускорение движения, повидимому, мы также не в состоянии определить с точностью; при более же быстром движении, в особенности при известной его форме, именно при вращении вокруг оси, мы начинаем испытывать головокружение, выражающееся ложным ощущением движения нашего тела.
1 «Путешествуя по железной дороге, — говорит Мах, — мы ощущаем только маленькие сотрясения, которые вызывают замедления или ускорения в движении нашего тела; но так как средняя скорость их остается неизменною при движении поезда вперед или назад, то эти сотрясения друг друга уравновешивают. И действительно, с закрытыми глазами мы можем легко представить себе то или другое движение поезда и даже без труда перейти от одного представления к другому. Последнее становится невозможным, когда поезд только что начинает двигаться или когда замедляет свой ход перед остановкой; в этом случае ускорение ие равно нулю и имеет определеннее направление.^ (Mach, 1/с, р. 23).
14Я
Таким образом, органы равновесия не могут считаться приспособленными к восприятию ощущений движения тела; они лишь воспринимают ускорение этого движения, а не самое движение, и лишь этим путем дают нам возможность определять направление движения.
То обстоятельство, что разрушение или угнетение функции того или другого органа равновесия на одной стороне, как мы видели, сопровождается ложным ощущением движения тела, еще ничуть не говорит в пользу гипотезы Маха. Дело в том, что органы равновесия, как уже я выяснил, при нормальных условиях находятся в непрерывно деятельном состоянии; при этом обычное покойное положение тела, например, вертикальное, предполагает почти одинаковые или тождественные ощущения от органов равновесия той и другой стороны. Всякая же перемена покойного положения вперед или назад, вправо или влево, а тем более вращение тела вокруг оси сопровождается уже усилением центростремительных импульсов в органах одной стороны и ослаблением их в органах другой стороны.
При одностороннем разрушении и вообще при угнетении функции одного из органов равновесия мы должны предполагать подобные же отношения: вместе с прекращением возникающих •в этом органе ощущений импульсы, посылаемые к центрам от органов равновесия другой стороны, не будут в достаточной мере уравновешиваться импульсами от неповрежденных органов равновесия на стороне разрушения, вследствие чего в этом случае должна произойти дисгармония в ощущениях, исходящих от органов равновесия той и другой стороны, совершенно такого же рода, как и при быстром движении в сторону и в особенности при вращении тела вокруг оси у здоровых животных.
Подобная же дисгармония в ощущениях должна произойти и при неполных двусторонних разрушениях одного из органов равновесия (перерезка передней или задней пары каналов, разрушение передней или задней стенки третьего желудочка и пр.). Естественно поэтому, что как одностороннее разрушение или вообще угнетение функции органов равновесия, так и двустороннее, но ограниченное их повреждение должны сопровождаться не чувством ненормального положения тела, а ложным ощущением движения, иначе говоря, ощущением непрерывной перемены положения тела. Напротив того, в случаях двусторонних и значительных разрушений органов равновесия мы должны предполагать более или менее полную утрату чувства положения головы и тела вообще. Таким образом, весь анализ фактов приводит нас к выводу, что такназываемыеперифе-рическиеорганы равновесия, являющиеся приводам имозжечк а, помимо их рефлектор-
144
ной функции, служат органами, восприн и мающими ощущения положения тела.
В пользу такой роли органов равновесия говорит, безусловно, и анализ явлений, наблюдаемых при головокружении, развивающемся как под влиянием вращения тела вокруг оси, так и при гальванизации в области затылочной части головы.
Мы уже видели выше, что явления головокружения в этих случаях зависят от нарушения функции органов равновесия. Мы выяснили, кроме того, что основное ощущение, возникающее под влиянием этого нарушения функции органов равновесия, именно субъективное чувство движения тела в известном направлении, переносится нами на внешний мир или объективируется, вследствие чего происходит кажущееся движение в противоположном направлении тех из окружающих предметов, которые мы видим, осязаем и звуки которых мы слышим; при этом само восприятие внешних впечатлений (зрительных, осязательных и слуховых) не страдает в значительной степени, за исключением только случаев сильного головокружения.
Но что же такое в действительности это кажущееся движение предметов окружающего мира, • как не постоянное нарушение проицирования наших специальных ощущений? Не значит ли это явление то, что мы в состоянии еще воспринимать внешние впечатления, которые возбуждают в нас зрительные, слуховые и осязательные ощущения, но что проекция этих ощущений представляется нарушенною? Иначе говоря, при головокружении вместо того, чтобы относить наши зрительные, слуховые и осязательные ощущения или образы в том направлении, в каком располагаются воспринимаемые нами объекты внешнего мира, мы относим эти ощущения в другие точки пространства, причем проекция их не представляется уже устойчивой, а как бы постоянно нарушается в известном направлении.
Так как все вообще органы равновесия функционируют одновременно, причем получаемые нами ощущения положения тела являются результатом их совместной деятельности, то естественно, что даже в том случае, когда мы имеем дело с поражением одного лишь органа равновесия (например, одних полукружных каналов), головокружение всегда является в виде нарушения проекции всех вообще пространственных ощущений (зрительных, слуховых и осязательных). С разрушением или поражением одного из этих органов, как было выяснено, нарушается гармония в доходящих до сознания импульсах, исходящих из органов равновесия, результатом чего, с одной стороны, являются рефлекторные движения, с другой — в нашем сознании возникает постоянное нарушение чувства положения тела и проекции наших ощущений в пространстве. Эти расстройства со временем, однако, могут выровняться благодаря тому, что неповрежденные органы равновесия принимают на себя функцию утраченного или поврежденного органа. В таком
10 В. М. Бехтерев
145
случае как двигательные расстройства, так и субъективные явления головокружения постепенно исчезают. Это и объясняет нам, почему в случаях старых поражений мозжечка и полукружных каналов головокружение иногда совершенно отсутствует.
Таким образом, в головокружении, зависящем от нарушения функции органов равновесия, мы встречаем самый поразительный пример восприятия наших ощущений, получаемых при посредстве специальных органов внешних чувств (зрения, слуха и осязания), с нарушением того именно процесса, который понимается под названием проекции ощущений и который лежит в основании всех наших пространственных представлений.
Но раз существует возможность изолированного нарушения проекции наших ощущений при нарушении функции органов равновесия, то очевидно, что процесс проицирования ощущений никак нельзя ставить в исключительную зависимость от функции специальных органов, которые служат для восприятия внешних впечатлений.
Проекция ощущений наружу, по нашему мнению, должна зависеть главнейшим образом отфункцииособых органов нервной системы, называем ыхнамиорганамиравно-в е с и я.
Это заключение само по себе представляет только неизбежный вывод из наблюдаемых фактов. Как на основании двигательного или чувствительного паралича, являющегося результатом повреждения определенных частей мозга, мы заключаем об отношении этих частей к функции движения или чувствительности, так и на основании нарушения в проекции ощущений при головокружении, составляющем последствие нарушения функции органов равновесия, мы заключаем, что эти последние органы играют выдающуюся роль в процессе отнесения наружу наших специальных ощущений.
Значение органов равновесия в отношении проицирования зрительных ощущений, между прочим, прекрасно выясняется на слепорожденных, подвергнутых операции. Известно, что такие лица уже с первых моментов после операции относят свои зрительные образы наружу или объективируют их, хотя и очень несовершенно, определяют форму, размеры и расстояние видимых ими предметов. Так как в этом случае воспри'ятию зрительных объектов не предшествовало никакого опыта со стороны органа зрения, то, очевидно, это обстоятельство говорит в пользу того, что в нашем организме должны существовать особые условия, благодаря которым сознание тотчас же убеждается, что дело идет о восприятии, источник которого находится вне нас. Эти-то особые условия и даны в ощущениях положения нашего тела, воспринимаемых органами равновесия.
Мы не говорили до сих пор о теоретических воззрениях Циона, близко относящихся к предмету нашего исследования,
146
только потому, что находили удобнее обсудить их вслед за указанными теориями.
Упомянутый автор в последней своей работе о полукружных каналах1 впервые довольно подробно изложил гипотезу об отношении этих органов к образованию наших представлений о пространстве. Он сам формулировал выводы этой части своей работы следующим образом: «полукружные каналы суть периферические органы пространственного чувства, т. е. ощущения, вызываемые раздражением нервных окончаний в ампулах, служат для образования наших понятий о трех измерениях пространства. Ощущения каждого каналасоответствуют одному из этих измерений. Помощью этих ощущений в нашем сознании образуется представление идеал ь-ногопространства, на котороепереносятся как все ощущения о расположении предметов в пространстве, доставляемые нам другимиорганами чувств, так и ощущения о положении нашего тела в пространств е».
Факты, на которых основывалась эта попытка объяснить образование наших представлений о пространстве, далеко не многочисленны и притом наиболее существенные из них, по моему мнению, не имеют того значения, которое приписывает им сам автор. Главнейшие основания, на которых покоится гипотеза Циона, им самим были выражены в следующих словах: «Раз несомненно, с одной стороны, что наши представления зависят прежде всего от бессознательных иннервационных ощущений или ощущений сокращения глазных мышц, с другой стороны,—* что каждое, даже минимальное, возбуждение полукружных каналов вызывает сокращение и иннервацию этих же мышц, то очевидно, что нервные центры, в которых оканчиваются нервные волокна, разветвляющиеся в полукружных каналах, должны со1 стоятьвинтимной связисглазодвигатель-ным центром, и что, следовательно, их возбуждение должно оказывать преобладающее влияние на образование наших пред-ставленийопространстве».
Хотя сам Цион, признавая это положение простым выра-1 жением фактов, считает его безупречным, тем не менее от внимательного читателя не может, однако, ускользнуть то обстоятельство, что если первая часть этого вывода, относящаяся до связи центральных окончаний нервных волокон, принадлежащих полукружным каналам, с глазодвигательным центром, в действительности не может подлежать сомнению,
1 Ц и о и, These pour le doctorat, Paris, 1878; Воеино-медицинский журнал,- 1879.
10*
147
то далеко нельзя сказать того же о второй части вывода. Трудно, в самом деле, понять, каким образом на основании связи центров для полукружных каналов с глазодвигательными нервами можно выводить заключение о влиянии каналов на образование представлений о пространстве.
Быть может, вследствие неубедительности представленных Ционом доводов в пользу своего взгляда или по каким-либо другим причинам его гипотеза не только не получила широкого распространения, но и упоминается только немногими из авторов, писавших о полукружных каналах позднее Циона.
Действительно, уже в основании своем эта гипотеза имеет одну слабую сторону, на которую я намерен здесь указать. Допуская прирожденную нам организацию для возникновения идеи о пространстве, гипотеза Циона в сущности вовсе не выясняет нам, каким образом полукружные каналы служат для восприятия пространства и создания пространственных представлений. Цион, правда, говорит в одном месте, что ощущения каждого канала соответствуют одному из трех измерений пространства; но какие это ощущения по существу и в каком отношении они соответствуют одному из трех измерений пространства —на это мы не находим ясного ответа.
С другой стороны, мы встречаем у Циона указание, что полукружные каналы представляют собою «специальный орган пространственного ощущения». По его воззрению, трудности, с которыми сталкиваются эмпирические и нативистические теории об образовании наших представлений о пространстве, «совершенно исчезают, если допустить существование органа чувств, специально предназначенного для того, чтобы доставлять ощущения, служащиепрямодля образов ания понятия о пространстве с тремя измерениям и». Подобный орган Цион и видит в системе полукружных каналов. Несколько ниже автор говорит: «Мы можем очень хорошо понять, каким образом рядволокон, расположенных в направлении трех измерений, дает нам при возбуждении ряд бессознательных ощущений о пространстве, имеющем подобные измерения». Однако вряд ли кто может согласиться с этими заключениями автора. Легко решить вековую задачу, сказав, что мы имеем «специальный орган пространственного ощущения», благодаря которому «в нашем сознании образуется представление идеального пространства», на которое переносятся все наши ощущения, но такой способ понимания весьма трудно помирить с нашими общими воззрениями об отправлениях нервной системы. Если бы мы, например, могли еще представить себе, что и другие органы чувств с самого уже начала дают в нашем сознании нечто готовое «идеальное», с чем бы только сравнивались остальные ощущения, то выше указан,-
148
ный взгляд еще мог бы иметь какую-нибудь почву под собою, а так как подобных органов мы не знаем, то самая сущность гипотезы Циона противоречит основным понятиям об отправлениях нервных центров вообще.
Помимо того, против гипотезы Циона может быть выставлено и множество других возражений. Если, в самом деле, в полукружных каналах мы обладаем органом пространственного чувства, при посредстве которого создается в нашем представлении идеальное пространство, на которое только переносятся ощущения, получаемые нами от органов внешних чувств, то как можно объяснить себе то обстоятельство, что одни ощущения мы локализируем во внешнем пространстве, другие же —нет?
Наконец, с чисто физиологической стороны гипотеза Циона не выдерживает критики уже потому, что в настоящее время мы знаем еще и другие органы, по функции совершенно сходные с полукружными каналами.
Высказывая эти замечания по поводу гипотезы Циона, мы, однако, ни на одну минуту не сомневаемся, что органы равновесия, действительно, играют известную роль в отношении образования наших представлений о пространстве, но роль их в этом отношении мы понимаем совершенно иначе. Прежде всего органы равновесия играют несомненную роль в том процессе, который может быть назван проекцией наших ощущений наружу и который лежит в основе всякого пространственного восприятия. В последнем, на наш взгляд, должно различать два процесса: а) проекцию ощущений наружу и б) локализацию их в окружающем пространстве. Первый процесс есть основной, так как на основании его впервые и создаются представления о «я», как субъекта, и «не я», или объекте, иначе говоря представления о внутреннем и внешнем мире. Без этого первоначального различения немыслима была бы и локализация ощущений в пространстве.
Объяснить закон проекции — значит объяснить первоначальное восприятие пространства. Большинство теорий локализации ощущений в пространстве предполагает проекцию ощущений как данную нам, между тем с нее собственно и следовало бы начинать исследование. В самом деле: почему мы воспринимаем ощущения в виде внешних образов, т. е. образов, относимых нами наружу, во внешний мир, а не в виде внутренних состояний, каковы, например, чувствования? Когда мы разрешим этот вопрос, то сделаем тем самым первый и существенный шаг в объяснении локализирования наших ощущений.
Проще всего, конечно, изучать явление на проекции предметов, доступных нашей руке. Представим себе, что мы осязаем данный предмет. Почему он кажется нам прикасающимся снаружи? Потому что, изменяя положение руки, мы изменяем и самое ощущение; мы отнимаем руку — оно исчезает, приближаем, и оно вновь является. Ясно, следовательно, что предмет
149
ощущения лежит вне нас. Следовательно, мы делаем здесь заключение, но это заключение возможно лишь благодаря ощущению положения своей руки. Подобную же роль должны играть и органы равновесия, дающие нам ощущения положения головы и тела. Очевидно, прежде чем относить ощущения наружу, мы должны иметь ощущение положения собственного тела. Без таких ощущений немыслимо и отнесение ощущений кнаружи от нас.
Но мы уже видели, что мы ощущаем с точностью положение своего тела при посредстве органов равновесия. Очевидно, таким образом, что, благодаря деятельности этих органов, мы и относим или проицируем наши ощущения наружу.
Так как органы равновесия даны нам от природы, то отсюда ясно, что закон проекций ощущений имеет для себя готовую органическую основу. С другой стороны, представляется более чем очевидным, что если мы имеем специальые органы, функция которых столь тесно связана с проекцией ощущений наружу, то эти же органы должны играть видную роль и в образовании наших представлений об окружающем пространстве.
Постоянно возбуждаясь при всех обыкновенных условиях и функционируя в общей гармонии друг с другом, органы равновесия, как мы видели, доводят до сознания в каждый данный момент ощущения положения головы и тела. Эти ощущения меняются с каждым перемещением нашего тела, благодаря чему это перемещение и воспринимается нами как таковое. Следовательно, благодаря нашим органам равновесия, мы непосредственно чувствуем, что мы двигаемся где-то, а это самое и служит нам первым шагом к образованию представлений об окружающем нас пространстве. Таким образом, с помощью непосредственно воспринимаемых нами ощущений положений головы и тела мы и с о-здаем понятие об окружающем нас пространстве с тремя измерениями, которое, согласно мнению Канта, является «необходимым предположением» и от которого сознание наше даже не может отрешиться ни на одну минуту.
В этом отношении черепные органы равновесия (полукружные каналы и область третьего желудочка) разделяют свою роль с тем сложным чувством, которое дает нам возможность отделять положение своих членов и которое, как мы видели, по всей вероятности, также должно быть сведено на функцию особых нервных аппаратов, которые мы называем кожно-мышечными органами равновесия. В самом деле: двигая своими членами вправо и влево, вверх и вниз, взад и вперед и получая каждый раз ощущения о положении своих членов, мы одинаковым образом убеждаемся, что вне нас имеется пространство, в котором движутся наши члены.
150
Трудно сказать, ощущения ли положения головы и тела, воспринимаемые нами главным образом при посредстве черепных органов равновесия, или же чувство положения членов играют более важную роль в отношении создания наших представлений о пространстве. Можно лишь сказать, что от первых органов мы получаем более интенсивные ощущения в том случае, когда наше тело испытывает массовое перемещение (например, при ходьбе), тогда как чувство положения членов дает нам особенно резкие ощущения при перемещении последних (например, при движениях рук. ног1. В известных случаях, как, например, при пассивном массовом передвижении тела, мы получаем ощущения перемещения тела только от черепных органов равновесия, но, с другой стороны, имеются также случаи, где мы получаем ощущения перемещения частей своего тела исключительно с помощью чувства положения членов, как, например, при простом передвижении глазных осей, при перемещении кисти или пальцев руки.
Очевидно, таким образом, что черепные органы равновесия и кожн о-м ышечные органы в отношении восприятия ощущений положения тела него членов взаимно до-полняютдругдругаислужатводинаковой мере к созданию наших представлений о пространстве.
В этом-то пространстве, т. е. в пространстве, созданном ощущениями положения тела и его членов, мы. и локализируем наши специальные ощущения, причем более точное размещение в пространстве зависит главным образом от соответствующих приспособлений в органах чувств и воспитания последних согласно учению эмпирической теории.
В чем же причина того, что наши ощущения мы неизбежным образом располагаем в пространстве по трем измерениям?
Когда общее представление пространства уже создалось на основании получаемых нами ощущений положения тела и его членов, то необходимость расположения в трех измерениях пространства наших специальных ощущений, получаемых нами от органов чувств, обусловливается уже тем, что вместе с перемещением нашего тела или его органов, следовательно, вместе с изменением ощущений нашего положения изменяется каждый раз и положение воспринимаемого впечатления в отношении соответствующего органа, а вместе с этим, следовательно, оно должно сопровождаться уже иными «местными знаками». Отсюда неизбежно устанавливается известное соотношение между ощущением положения нашего тела и его членов и определенным изменением местных знаков в ощущениях, воспринимаемых органами внешних чувств, а это в свою очередь дает сознанию ясное указание о направлении источника внешнего раздражения по отношению к нам самим. Следовательно, в соотношении между ощущениями положен ия с о б-
Г5Г
ственноготелаиегочленовиместнымиз на-ками в ощущениях и заключается причина того, почему мы располагаем свои ощущения по трем измерениям пространства.
Мы знаем, однако, что наши ощущения не только размещаются нами в окружающем пространстве по трем измерениям, т. е. относятся во внешний мир по тому или иному направлению, но некоторые из ощущений, как, например, зрительные и осязательные, мы локализируем также в известных пунктах окружающего нас пространства и можем в то же время определить с известной точностью как расстояния внешних предметов от нас самих, так и размеры этих предметов.
В этой части пространственного восприятия, кроме местных знаков, наиболее деятельную роль играет прежде всего чувство положения членов; но, очевидно, также и черепные органы равновесия принимают здесь некоторое участие.
Перемещаясь в пространстве и протягивая руку к окружающим предметам, мы получаем возможность на основании воспринятых нами ощущений определить расстояние того или другого из предметов до нас самих, следовательно, можем локализировать его в известной части окружающего пространства. С другой стороны, на основании ощущений о положении глаз в орбитах, дающих нам понятие о степени сведения глазных осей, и на основании напряжения аккомодации мы можем локализировать предметы в известном расстоянии от нас самих и не приближаясь к ним.
Но, как известно, и при неподвижном положении нашего тела и наших глазных осей мы до известной степени оцениваем относительное положение прикасающихся к нам и видимых нами предметов по тем местным знакам в соответствующих ощущениях, значение которых так прекрасно оценил Гельмгольц. Эти местные знаки играют, очевидно, также известную роль и в определении размеров окружающих предметов, в чем, впрочем, немаловажное значение должно принадлежать также чувству положения членов (обхватывание предметов, перемещение глазных осей на различные части предмета и т. п.), а также ощущениям положения головы и тела, воспринимаемым при посредстве черепных органов равновесия (перемещение головы и туловища при осматривании и прикосновении к различным частям предметов). Утрата чувства положения членов в пространстве лишает нас не только возможности оценивать размеры схватываемых нами предметов, но и возможности оценивать расстояние от нас и относительное положение по отношению к нам тех предметов, до которых мы касаемся. Точно так же и утрата того или другого из черепных органов равновесия лишает нас не только ощущения прочности нашего положения, но, как мы видели, сопровождается также нарушением проекции наших ощущений в пространстве, выражающемся сильным головокружением.
152'
Когда последнее достигает наибольшей степени развития, то мы временно утрачиваем даже способность объективировать наши ощущения, появляется потемнение в глазах и дело может дойти до помрачения сознания.
Таким образом, ощущения, получаемые нами от черепных органов равновесия, кожно-мышечные ощущения, в основе чувства положения членов, и местные различия в ощущениях, получаемые при раздражении различных частей того или другого из органовчувств, илитак называемые местные знаки, суть главнейшие виновники нашего пространственного восприятия.
Справедливость этого заключения, между прочим, выясняется и из того, что все те ощущения, как, например, центральные чувства, которые по самому характеру своему не вступают в постоянное соотношение с ощущениями положения тела и его членоч и в то же время лишены местных знаков, не могут быть относимы к внешнему пространству. Они суть вполне субъективные ощущения, лишенные всякой локализации даже и в нас самих, если не считать за таковую локализирование в определенных частях тела их периферических соощущений (чувство сжатия и стеснения в сердечной области при тоске и т. п.).
Уже в начале работы мы обращали внимание, между прочим, на близкое отношение периферических органов равновесия к высшим органам чувств (полукружных каналов — к органу слуха, области третьего желудочка — к органу зрения, кожно-мышечных органов — к чувству прикосновения и давления), что говорит с вероятностью в пользу функциональной связи между теми и другими. Существование такой связи выясняется в особенности из того обстоятельства, что впечатления, воспринимаемые тремя высшими органами внешних чувств, resp. зрительные, слуховые и осязательные импульсы, нередко оказывают прямое влияние на функцию равновесия. Последний факт в свою очередь может быть объяснен или путем непосредственной передачи упомянутых импульсов к соответствующим органам равновесия, или одновременным раздражением этих органов. В том и другом случае наши впечатления, служа, так сказать, внешними раздражителями органов равновесия, в свою очередь должны возбуждать в последних соответствующие ощущения.
Возможно и даже вероятно, что и эти ощущения, возбуждаемые при посредстве внешних раздражений органов равновесия, не остаются без значения в отношении локализации соответствующих им специальных ощущений, но во всяком случае вопрос этот нуждается еще в специальном исследовании.
О ТАК НАЗЫВАЕМОМ СУДОРОЖНОМ ЦЕНТРЕ
И О ЦЕНТРЕ ПЕРЕДВИЖЕНИЯ ТЕЛА
НА УРОВНЕ ВАРОЛИЕВА МОСТА
Как известно, до настоящего времени большинством авторов признается существование особого «судорожного» пентра на уровне варолиева моста, которому.приписывают появление судорог при усиленной венозности крови (как, например, при задушении), под влиянием внезапной анемии тела (например, при быстрой перевязке обеих сонных и подключичных артерий, а также при обширных кровотечениях), наконец, при быстром венозном застое (например, вследствие перевязки выходящих из головы вен). Принимают обыкновенно, что этот же судорожный центр приходит в возбуждение при действии некоторых ядов, например, сердечных, а также при действии никотина, пикротоксина и пр. В пользу существования этого центра приводили также тот факт, что непосредственное раздражение продолговатого мозга химическими реагентами, электричеством и механическим путем вызывает общие судорожные явления. Возбуждению этого же центра приписывалось и развитие общих судорог в приступе падучей.
Учение о «судорожном» центре на уровне варолиева моста получило особенное развитие со времени исследования Нотна-геля, который в своих опытах над кроликами убедился, что путем вкола в область варолиева моста удается вызвать настоящий падучевидный приступ. При своих исследованиях этот автор пытался определить также границы описываемого им «судорожного» центра на уровне варолиева моста. По этим исследованиям, судорожный центр помещается в области дна четвертого желудочка над ala cinerea и простирается вверх до четверохолмия; внутреннюю его границу составляют eminentia teres; наружная определяется труднее, но она не заходит за locus cinereus и tub. acusticum.
В прежнее время, когда мы еще не имели прочных и вполне доказательных сведений о важном участии двигательной области мозговой коры в развитии судорог в течение падучного приступа, естественно было относить развитие этих судорог на счет возбуждения своеобразного «судорожного» центра, значение кото-
154
рого в организме, помимо участия в развитии падучевидных судорог, представлялось совершенно загадочным. В новейшее же время с выяснением важной роли двигательных областей мозговой коры в развитии падучевидных судорог возникает, естественно, вопрос, как должно смотреть на те падучевидные судороги, которые развиваются при раздражении судорожного центра?
Уже многие авторы, признававшие особо важное значение двигательных областей мозговой коры в развитии падучевидных судорог, выражали сомнение в том, что судороги, наблюдаемые при раздражении основных частей мозга, а следовательно, и области так называемого судорожного центра, имеют действительно падучевидный характер. Наблюдения над кроликами представлялись, между прочим, недостаточными потому, что у этих животных будто бы не удается вызвать даже и с мозговой коры типичного падучного приступа. Исследования же, производимые с этой целью на собаках, у которых производилось непосредственное раздражение области продолговатого мозга, как будто говорили в пользу упомянутого сомнения, так как вышеуказанным путем у них вызывались лишь общие тонические судороги, прекращавшиеся тотчас же вслед за прекращением раздражения. Эти данные, таким образом, как бы разрешали вопрос в том смысле, что раздражение судорожного центра само по себе не может дать настоящего падучного приступа. При этом существование самого судорожного центра не подвергалось сомнению, а исключалось лишь его участие в развитии судорог, наблюдаемых в течение падучного приступа.
Исследования, произведенные в заведываемой мною лаборатории первоначально доктором Сущинским, а затем доктором Вырубовым, которым они, собственно, и были доведены до конца, разъясняют самый вопрос по существу. Эти исследования подтверждают вполне тот факт, что путем вкола иглы в область варолиева моста у кролика вызывается настоящий падучевидный приступ. По описанию доктора Вырубова, вслед за указанным вколом у кролика тотчас же развивается приступ общих судорог, выражающихся первоначально резким спинным столбняком (opisthotonus) и тоническим вытяжением мышц конечностей, после чего развиваются клонические судороги, распространяющиеся на мышцы большей части тела. При этом глазные яблоки стоят неподвижно, роговица становится нечувствительной, зрачки расширяются и не реагируют на свет. Вместе с этим потухают совершенно или по крайней мере резко понижаются и другие рефлексы, как, например, коленные, которые, согласно ранее сделанным Говерсом и мною наблюдениям, а также наблюдениям доктора Васильева (из моей клиники), совершенно прекращаются или резко понижаются непосредственно вслед за падуч-ными приступами. Словом, у кроликов после вкола в варолисв мост существовали налицо все характерные признаки падучного
155
приступа, и есть полное основание думать, что и сознание при этом утрачивалось; в пользу этого говорит то обстоятельство, что при вышеуказанных приступах выражение лица животного становилось как бы безжизненным, маскообразным.
Таким образом, не подлежит сомнению, что у кроликов механическим раздражением области варолисва моста удается вызвать настоящий падучевидныи приступ, ничем по существу не отличающийся от обыкновенного падучного приступа.
Неоднократно присутствуя при опытах, производимых докторами Сущинским и Вырубовым, я могу с положительностью утверждать, что вызванные вышеуказанным образом судорожные приступы совершенно аналогичны по своим проявлениям и течению падучным приступам, вызванным у тех же животных иным образом, например, раздражением мозговой коры, с помощью которого, вопреки утверждению некоторых авторов, а у кроликов, подобно другим животным, могут быть вызваны настоящие падучные приступы. Но если даже иметь в виду сравнение вызванных у кролика вколом в область варолиева моста судорожных приступов с падучными приступами, например, собаки, то и в таком случае нельзя провести существенного различия между теми и другими приступами.
Надо, впрочем, заметить, что и у собак, у которых простой вкол в варолиев мост обычно не дает падучного приступа, электрическим раздражением области варолиева моста, произведенным путем вкола в него нескольких игл и соединения их с электродами от катушки Дюбуа-Реймона, доктору Вырубову удавалось вызывать настоящие падучные приступы. Ясно, следовательно, что нельзя более говорить о том, что путем раздражения варолиева моста у собак не могут быть вызваны падучные приступы, как это утверждали некоторые авторы.
Возникает, однако, вопрос, чем обусловливаются эти падучные приступы, которые с таким постоянством наблюдаются у кроликов при вколе в pons Varolii. Действительно ли здесь дело идет о раздражении особого «судорожного» центра и не участвуют ли в происхождении этих приступов двигательные области мозговой коры? Опыты, произведенные с этой целью в заведываемой мною лаборатории доктором Вырубовым, вполне разъясняют этот вопрос. Оказывается, что если предварительно животному произвести перерезку на уровне четверохолмия, то у него развивается резкий спинной столбняк (opisthotonus) и быстро исчезающие клонические судорожные подергивания.
Если вслед за тем кролику произвести вкол в область варолиева моста, то в этом случае весь эффект выражается лишь некоторым усилением спинного столбняка, клонических же судорог уже не наблюдается; иначе говоря, при данных условиях падучных приступов не наступает.
Равным образом, если у кролика предварительно удалить мозговые полушария, сохранив в целости узлы основания мозга,
156
то и в таком случае вкол в область варолиева моста не вызывает падучного приступа; появляется лишь несколько разбросанных судорожных подергиваний.
Из этих данных вытекает с несомненностью, что падучные или падучевидные приступы, развивающиеся у животных при раздражении области варолиева моста, ничуть не обусловливаются раздражением лежащего здесь «судорожного» центра, а вызываются благодаря передаче этого раздражения на мозговые полушария и, в частности, на двигательные области мозговой коры, что, собственно, и служит причиной развития падуче-видных приступов при раздражении области варолиева моста.
Другой ряд исследований с целью выяснения того же вопроса был произведен в заведываемой мною лаборатории доктором И. К- Мейером над токсической падучей, вызванной отравлением с помощью впрыскивания животным абсента и цинхонина.
Если у собак или кошек вызвать с помощью вышеуказанных средств падучный приступ и затем быстро, в момент самого приступа, произвести перерезку на уровне четверохолмия, то вслед за тем тотчас же клонические судороги прекращаются, и у животных остается лишь общий спинной столбняк и вытяжение конечностей, т. е. явления, которые вызываются операцией перерезки и без предварительного вызывания падучного приступа. С другой стороны, у животных, которым предварительно была произведена перерезка на уровне четверохолмия, а равно и в тех случаях, когда производилось полное и двустороннее удаление двигательных областей мозговой коры, введением вышеуказанных средств в кровь животных уже не удавалось вызвать настоящего падучного приступа.
Таким образом, эти исследования стоят в полном согласии с вышеизложенными данными и также, как и эти последние, не оставляют сомнения в том, что развитие судорог в течение падучных приступов обусловливается главным образом участием мозговой коры; основные же части мозга если и принимают какое-либо участие в развитии падучевидного приступа, то лишь в той его части, которая выражается развитием тонических судорог.
Спрашивается теперь, чем объяснить передачу раздражения с области варолиева моста на мозговую кору, когда вызывается падучный приступ вколом иглы в область варолиева моста? Ввиду того что произведенные в заведываемой мною лаборатории опыты доктора Тодорского и в последнее время доктора Боришпольского доказывают с несомненностью, вопреки взгляду других авторов, что в основе падучных приступов — будут ли они вызваны введением яда в кровь (например, абсента) или же электрическим раздражением мозговой коры — всегда лежат явления прилива крови к мозгу при активном сужении артериальных сосудов тела, я склонен думать, что при уколе в варолиев мост у кроликов дело идет также о приливе крови к мозгу при
157
активном сужении артериальных сосудов в остальных частях тела, что и обусловливает развитие падучного приступа.
Если у собаки не удается достичь того же результата с помощью простого вкола в варолиев мост, то это объясняется, по моему мнению, лишь тем, что у более высших животных, благодаря большим размерам мозга, простым вколом в варолиев мост нельзя привести в возбуждение сосудодвигательный центр; при более же значительном повреждении вовлекаются в последнее и двигательные проводники, что препятствует само по себе развитию падучных приступов. Ввиду этого и требуется у таких животных применение совершенно специального способа электрического раздражения области варолиева моста для вызывания падучного приступа, как делал в заведываемой мною лаборатории доктор Вырубов в отношении собак.
Но как бы ни смотреть на способ передачи раздражения с варолиева моста на область мозговой коры, не подлежит сомнению, что вышеуказанные опыты делают существование судорожного центра варолиева моста совершенно невероятным. В самом деле, какую же роль должен исполнять судорожный центр в организме, если даже общие падучевидные судороги зависят не от раздражения этого центра, а от раздражения мозговой коры? Ясно, что с этим вместе утрачивается наиболее веский довод в пользу существования особого судорожного центра на уровне варолиева моста.
Само собою разумеется, что этим мы ничуть не хотим сказать, что в основных частях мозга и, в частности, в области варолиева моста, не содержится вовсе двигательных элементов. Напротив того, мы считаем несомненным, что такие элементы существуют, но признавать их за особый судорожный центр было бы также неправильно, как неправильно называть судорожными центрами двигательные клетки передних рогов спинного мозга.
Не подлежит никакому сомнению, что на уровне верхних частей продолговатого мозга и варолиева моста имеется особый двигательный центр, но он представляет собою не судорожный центр, а центр передвижения. В самом деле, существование такого центра может быть доказано уже у лягушек.
Последние, как известно, могут еще ползать и прыгать даже и после удаления мозжечка, но лишь в том случае, если у них сохранена верхняя треть продолговатого мозга; с разрушением же последней лягушки совершенно лишаются способности передвижения. По Тарханову, даже в том случае, если у лягушки мы удалим вместе с соответствующей половиной мозжечка прилежащий отдел верхней части продолговатого мозга, животное еще не утрачивает способности ползать; последнее обстоятельство доказывает существование в этих областях мозга парного сочетательного механизма передвижения, причем от каждой половины этого механизма отходят нервные проводники ко всем четырем конечностям. Следует, однако, заметить, что опериро-
158
ванные вышеуказанным образом лягушки могут лишь ползать, но не могут производить прыжков, для которых, очевидно, требуется участие обеих половин сочетательного механизма передвижения. С другой стороны, определенными разрезами, нарушающими связь перекрестных путей в спинном мозгу, можно лишить лягушку, у которой верхние части продолговатого мозга сохранены, способности ползания, тогда как она будет еще в состоянии производить прыжки (Тарханов). Ясно, что последние хотя и требуют участия обоих парных центров передвижения, но могут происходить лишь при сохранении прямой их связи с нижележащими спинномозговыми центрами конечностей, тогда как ползание может происходить и при сохранении в целости одного из центров передвижения; но зато необходимым условием для этого движения является сохранение перекрестных связей в спинном мозгу.
Не подлежит никакому сомнению, что и у высших животных имеется такой же центр передвижения, как и у лягушек, на уровне варолиева моста. И здесь удаление всех частей мозга до уровня моста хотя и лишает животное способности стоять вследствие нарушения равновесия тела, благодаря отделению области третьего желудочка, содержащей особый орган равновесия1, но при всем том у животного еще сохраняется способность правильного сочетаннсго движения конечностей, как при беге под влиянием болевых раздражений. Между тем при перерезке на уровне продолговатого мозга вышеуказанная способность совершенно утрачивается. Таким образом, приблизительно в тех частях мозга, в которых признается существование так называемого «судорожного» центра, мы имеем, несомненно, особый парный центр для передвижения тела, который приводит в сочетательную деятельность все конечности животного.
Очевидно, что раздражением этого двигательного центра для перемещения животного и могут обусловливаться судорожные движения, развивающиеся после сильных кровотечений при перевязке всех четырех мозговых артерий и при перевязке мозговых вен даже у тех животных, которым предварительно были удалены все части мозга впереди варолиева моста. Но ввиду вышеприведенных опытов Вырубова и Мейера не подлежит сомнению, что возбуждение этого центра не может дать в результате падучевидных судорог, выражающихся правильной сменой общих тонических судорог общими же клоническими мышечными сокращениями. Самое большее, этот центр может принимать участие лишь в развитии тонического периода падучного приступа, а ничуть не в развитии мелких клонических судорог, являющихся последствием ненормального возбуждения двигательных областей мозговой коры.
1 См. мою работу в Военно-медицинском журнале, 1883, и в Pfluger's Archiv, 1883.
159
В согласии с этим стоит, без сомнения, и тот факт, что раздражение области варолиева моста, как и двигательных путей белого вещества других мозговых областей, вызывает всегда лишь тонические, а не клонические судороги. Этим я не хочу сказать, что клонические судороги обусловливаются везде и всюду исключительно лишь раздражением мозговой коры. Напротив того, я убедился, что большие размашистые судорожные движения членов с характером клонических судорог могут происходить под влиянием сотрясения мозга даже у животных, которым произведено удаление двигательных областей мозговой коры. Точно так же под влиянием раздражения внекорковых областей могут происходить и отдельные судорожные подергивания, как это доказывают случаи хореи у собак, не прекращающейся даже и после перерезки верхних отделов спинного мозга. Но мелкая клоническая судорога, столь характеристичная для возбуждения двигательных областей мозговой коры, без сомнения, не может быть вызвана ни раздражением варолиева моста, ни раздражением других подкорковых областей.
О ВОЗБУДИМОСТИ МОЗГОВОЙ КОРЫ
у новорожденных животных
Вопрос о возбудимости двигательных центров у новорожденных животных стал интересовать ученый мир со времени исследований Зольтмана1, который убедился в том, что у щенят они представляются вначале совершенно не возбудимыми и только с 10-го дня по рождении впервые удается получить раздражением коры у щенят двигательный эффект в передней конечности, а с 13-го дня появляются уже движения в задних конечностях; к 16-му же дню центры лица и конечностей представляются уже вполне сформировавшимися. Исследования эти в отношении позднего развития двигательных центров коры у щенят и некоторых других новорожденных животных, например, кроликов и кошек, были затем подтверждены проф. Тархановым2 и мною3. Первый к наблюдению Зольтмана присоединил тот факт, что кроме животных, родящихся с недоразвитыми двигательными центрами, имеются животные, как, например, морские свинки, обладающие хорошо развитыми двигательными центрами уже при самом рождении. Недоразвитие центров у первых животных, по Тарханову, должно стоять в зависимости от недоразвития волокон пирамидного пучка и пирамидальных клеток мозговой коры, а не от недоразвития органов чувств, например, зрения, как предполагал Зольтман. Мои наблюдения показали, что развитие корковых двигательных центров у щенят происходит далеко не в одинаковое время. У одних —возбудимость центров впервые обнаруживается уже на 10-й день, у других же двигательные центры представляются невозбудимыми до 12 — 14-го и даже до 15-го дня; при этом я убедился также, что недоразвитие двигательных центров коры ничуть не находится в прямой зависимости от органов зрения, так как не существует никакого соотношения между открытием глаз животного и развитием психомоторных центров; с другой стороны, я убедился, что в то время как на поверхности мозга взрослых животных мы имеем
1 Jahrb. f. Kinderheilk., 1876, 9.
2 О психомоторных центрах у новорожденных животных и развитие их под влиянием разных условий, СПБ, 1879; Arch, slaves de biol., 1886.
» Врач, 1886, № 34.
И В. М. Бехтерев
161
значительное количество строго диференцированных центров, вызывающих сокращения определенных мышечных групп, у новорожденных щенят в первое время мы находим ограниченное число возбудимых центров, причем раздражение каждого из них вызывает общее сокращение мышц соответствующего члена, а не отдельных мышечных групп. Лишь постепенно, с течением времени происходит диференцирование двигательных центров для отдельных мышечных групп, участвующих в тех или других движениях. Далее я убедился, что с коры новорожденных животных до известного возраста нельзя вызвать клонической судороги членов, столь характеристичной для раздражения коры взрослых животных, а получаются всегда лишь медленные и одноразовые сокращения целого члена. Из других явлений заслуживает внимания отмеченное мною последовательное нарастание возбудимости двигательных центров, начиная с первого их появления, и поразительная истощаемость, обыкновенно тем большая, чем моложе животное, и, наконец, полная невозможность до известного времени вызвать у них падучные приступы. Надо заметить, однако, что наблюдения Панета1, Маркаччи2 и Ле-муана3, также проверявших исследование Зольтмана, говорят в пользу того, что возбудимость двигательных центров мозговой коры у большинства щенят появляется сравнительно рано. Лемуан, исследуя 2 новорожденных щенят и 2 новорожденных котят, убедился даже, что эффект раздражения коры у них был тот же салый, что и у взрослых животных.
Ввиду этих разноречивых данных я предложил работающему в моей лаборатории доктору Бари заняться систематическим исследованием вопроса о развитии двигательных центров мозговой коры у новорожденных животных.
Сделав до сего времени большое число опытов преимущественно над новорожденными щенками и котятами, он убедился, что у новорожденных щенят в возрасте до 20 дней из 38 опытов положительный результат раздражения корковых центров был получен в 25 случаях, в 12 же случаях, т. е. приблизительно в 1/3 случаев, был получен отрицательный результат; при этом все отрицательные опыты приходились на возраст до 9 суток. Отрицательный результат, по его мнению, мог зависеть в известных случаях от мозговой грыжи, кровоизлияния или охлаждения мозговой поверхности, но тем не менее в 4 опытах эффект получился отрицательный, несмотря на все предосторожности при опытах. Что касается опытов с положительным результатом, то в отдельных случаях удавалось получать сокращения членов даже у щенят в возрасте одних суток. При этом считаю необходимым заметить, что некоторые из этих опытов были произве-
1 Arch. i. d. ges. Phys., Bd. XXXVII, S. 203.
2 Marcacci, Giorn. d. R. Acad, di Torino, 1882.
3 These de Paris, 1880.
162
дены при моем личном участии. Сравнивая эти результаты с ранее произведенными мною исследованиями, приходится признать, что возбудимость мозговой коры у новорожденных животных находится в зависимости от самых разнообразных, нередко даже случайных условий, между которыми вместе с большей или меньшей зрелостью новорожденного животного, следует, вероятно, иметь в виду, между прочим, также индивидуальные отклонения.
Таким образом, разноречия во взглядах авторов относительно времени появления возбудимости корковых двигательных центров у новорожденных щенят получают достаточное объяснение в непостоянстве упомянутых результатов раздражения коры у животных одного и того же вида, как это было выяснено выше.
Далее доктору Бари при своих исследованиях над новорожденными животными удалось показать, что, кроме медленного, вялого характера сокращения членов, получаемого раздражением мозговой коры, скрытый период коркового мышечного сокращения у новорожденных животных представляется значительно длиннее скрытого периода коркового мышечного сокращения взрослых и что у новорожденных животных в противоположность взрослым разница между скрытым периодом коркового мышечного сокращения и мышечного сокращения, вызываемого раздражением подкорковых областей, представляется ничтожной.
Ввиду вышеизложенных результатов необходимо притти к выводу, что безмякотные недоразвитые волокна пирамидного пучка, повидимому, не вполне лишены возбудимости, но они, очевидно, лишены возможности передавать изолированные возбуждения к мышцам, как это мы наблюдаем у более развитых животьых, пирамидные волокна которых уже обложены мякотью.
11*
О РЕПРОДУКТИВНОЙ И СОЧЕТАТЕЛЬНОЙ РЕАКЦИИ
В ДВИЖЕНИЯХ
В мае 1907 г. вместе с осуществлением мною особого чувствительного дыхательного прибора с воздушно-водяной передачей явилась возможность исследовать влияние различного рода привычных сочетаний на такие отправления, как дыхание.
В работе, сделанной совместно с доктором Спиртовым, мы показали, что если приучить собаку сидеть спокойно в станке в темной комнате и начать производить звуковые раздражения, возбуждающие вдыхательную реакцию, то вслед за тем, как этот звук мы будем производить известное числе раз совместно со светом, не возбуждающим вдыхательной реакции, мы уже после нескольких повторений будем получать вдыхательную реакцию на один свет, не возбуждавший ранее вдыхательной реакции. Эта вдыхательная реакция после нескольких повторений одного светового раздражения исчезает, но достаточно вновь дать несколько совместных раздражений звука со светом, чтобы вызвать вновь ту же реакцию на один свет. Нет надобности говорить, что вышеуказанный способ дает возможность изучить отношения вдыхательной реакции при всевозможных взаимных отношениях звукового раздражителя со световым.
Аналогичные явления можно получить, взяв за основной раздражитель кожное болевое раздражение, например, от электрического тока, тогда как совместным раздражителем может быть взят по желанию световой, звуковой или какой-либо иной. В дальнейшем при исследованиях доктора Протопопова (в нашей лаборатории) выяснилось, что, если вызывать у собак вдыхательную реакцию с помощью кожного электрического раздражения, совмещая его с тем или другим тоновым раздражением, то оказывается, что не только при одном тоновом раздражении, но и подставляя вместо него любой тон, мы можем получать совершенно одинаковый вдыхательный эффект.
Отсюда очевидно, что по отношению к тоновой реакции со стороны дыхания не существует никакой специфичности. Факт этот заслуживает особого внимания ввиду того, что, как известно, по исследованиям доктора Зеленого над слюноотделительными условными рефлексами из лаборатории И. П. Павлова, дока-
164
зывалась специфичность реакции собак по отношению к тонам любого числа колебаний.
Ясно, что специфичность по отношению к одной из реакций организма ничуть не обусловливает подобной же специфичности по отношению к какой-либо другой реакции. Отсюда очевидно, что и отсутствие специфичности на определенную реакцию по отношению к одним раздражителям (например, цветовым раздражителям на слюноотделение у собак) * ничуть не исключает существования специфичности на какую-либо иную реакцию по отношению к тем же раздражителям.
Аналогичные результаты по отношению к вызыванию сочетательной реакции на индиферентное кожное раздражение после того, как оно совмещалось со звуком (выстрел), получены в нашей лаборатории в одном случае и на человеке (доктор Анфимов).
Имея в виду вышеуказанные результаты в отношении дыхательной функции, казалось вполне естественным перенести исследование и на обыкновенную двигательную реакцию у людей.
Для этой цели мы обратились к сгибательному движению положенного на резиновый баллон пальца у человека, которое регистрировалось на вращающемся закопченном барабане, благодаря воздушной передаче. Самое же исследование велось таким образом, что предварительно условливались с испытуемым, чтобы он реагировал сгибательным движением пальцев всегда лишь на данный звук, повторяемый с известной скоростью друг за другом, и после того, как он привыкал кэтой реакции, прекращали звук через тот или другой промежуток времени.
Оказывается, что при известной скорости следования звуковых раздражений друг за другом после внезапного прекращения их мы получаем еще в течение некоторого времени репродуктивную двигательную реакцию со скоростью следования бывших звуков, но постепенно ослабевающую.
При этом, чем чаще давался звук, тем большее число воспроизведений двигательной реакции наступает после прекращения звука. С другой стороны, число воспроизведений стоит в прямом соотношении с количеством произведенных звуков и с интенсивностью звуковых раздражений.
Таким образом, число воспроизведений двигательной реакции стоит в прямой зависимости от скорости следования бывших звуковых раздражений, от интенсивности взятого звука и от числа предварительно произведенных звуковых раздражений.
Само собою разумеется, что при работах этого рода мы не можем не встретиться с индивидуальными особенностями орга- • низма, но во всяком случае заслуживает внимания известная законность, о которой сказано выше.
1 О р б е л и, Доклад в Обществе русских врачей, 1907.
165
Подобные опыты ныне производятся в нашей лаборатории слушательницей Женского медицинского института Васильевой и результаты их будут сообщены впоследствии подробно.
С другой стороны, в работе слушательниц Женского медицинского института Нерпин и Добротворской имелось в виду выяснить, как такого же рода двигательная реакция у людей обнаруживается при замещении последовательного ряда звуковых раздражений, на которые должна происходить двигательная реакция, рядом раздражений иного рода, например, световых.
При этом в опытах, произведенных Добротворской, имелось в виду три случая:
1. В одном ряде опытов вместе с повторяемым через известные промежутки времени основным звуковым раздражением, на которое испытуемый должен был реагировать, давался одновременно и свет от электрической лампочки, на который реакции не требовалось и который время от времени оставлялся один, без звукового раздражителя, но со скоростью последнего.
2. В другом ряде опытов при повторении через известные промежутки времени основного звукового раздражения сопут-ствуемый ему свет давался не одновременно со звуковым, а всегда предшествовал ему за некоторое время, причем, как и в предыдущем случае, световой раздражитель время от времени оставлялся один, без звукового раздражителя.
3. В третьем ряде опытов при повторении через известные промежутки времени основного звукового раздражения каждый раз предварительно давался свет, который длился вплоть до наступления звукового раздражения, причем и здесь время от времени оставлялся один световой раздражитель.
При всех опытах в виде звукового раздражителя был взят обыкновенный электрический звонок, а для светового раздражителя пользовались обыкновенной же электрической лампочкой в 25 свечей, устроенной на уровне глаз испытуемого. Отметка испытуемыми производилась всегда сгибанием пальца правой руки.
Опыты делались в одних случаях при сосредоточении испытуемого на основном звуковом раздражителе, в других случаях при отвлечении от него посторонними раздражителями. Для последней цели на тыльной стороне предплечья левой руки писались волосяной кисточкой различные фигуры, за которыми испытуемый должен был следить, называя рисуемые фигуры.
В иных опытах для отвлечения испытуемого пользовались другим приемом; ему давали три трехзначных числа, причем к первому заставляли его присоединять по единице, из последнего отнимать по единице, а ко второму снова прибавлять по единице.
При данных опытах имелось в виду выяснить как скорость образования, так и стойкость сочетательной двигательной реак-
166
ции, в зависимости от силы света, продолжительности его как раздражителя, продолжительности антрактов между светом и звуком, их равномерности или неравномерности при повторениях, скорости следования звуковых раздражений друг за другом, количества данных комбинаций обоих раздражителей, навыка к движениям и тех или других индивидуальных особенностей.
Все опыты производились при одинаковых условиях, причем испытуемые во время опытов помещались в темной комнате. В'общей сложности слушательницей Добротворской было произведено 145 опытов над 3 испытуемыми.
Результаты этих опытов пока дают возможность сделать следующие заключения:
1. Появлению сочетательных двигательных реакций благоприятствует число произведенных опытов, благодаря чему с нарастанием числа опытов количество сочетательных реакций на один свет увеличивается.
2. Эти сочетательные реакции при повторении одного светового раздражителя постепенно слабеют, прекращаясь совершенно то раньше, то позже. Нередко угасание сочетательной реакции является лишь временным, так как та же реакция может появиться вновь без всяких внешних поводов, при дальнейших световых раздражениях.
3. Для возобновления угасшей сочетательной реакции требуется новое комбинирование прежних раздражителей, которое числом может быть менее числа комбинаций, вызвавших первоначальную сочетательную реакцию на свет.
4. Интенсивность света благоприятствует развитию и обнаружению сочетательных двигательных реакций.
5. Большая частота следования комбинированных световых и звуковых раздражений друг за другом также благоприятствует развитию и обнаружению сочетательных двигательных реакций на свет.
6. Равным образом и абсолютное количество комбинированных раздражений стоит в прямом отношении к развитию и обнаружению сочетательных двигательных реакций.
7. Из всех вышеуказанных комбинаций наиболее благоприт-ным для развития и обнаружения сочетательных двигательных реакций на свет является тот случай, когда свет предшествует звуку за две секунды, тогда как менее благоприятным оказалась комбинация с одновременным раздражением света и звука.
8. В случае предшествующего длящегося до звукового раздражения света оказалось, что как слишком большая длительность света, например, в 10—15 секунд, так и незначительная его длительность, например, в 2 секунды, представляются менее благоприятными условиями для развития и обнаружения сочетательных двигательных реакций, нежели промежуточная между вышеуказанными пределами длительность света.
167
9. Предварительное приучение к производству двигательной реакции вообще благоприятствует развитию и обнаружению сочетательных двигательных реакций.
10. Отвлечение испытуемого другой работой также благоприятствует образованию сочетательных двигательных реакций.
11. Неравномерность антрактов между раздражениями в ряде опытов была неблагоприятным условием для развития и обнаружения сочетательных двигательных реакций. Но у одного из испытуемых эта неравномерность явилась возбуждающим моментом по отношению к развитию и образованию сочетательных двигательных реакций.
12. Индивидуальность при вышеуказанных исследованиях играет большую роль. Прежде всего минимум необходимых комбинаций для образования сочетательных двигательных реакций и для их проявления зависит в значительной мере от индивидуальных условий. С другой стороны, и большая или меньшая сила сочетательных двигательных реакций, помимо всех вышеуказанных условий, находится в зависимости от индивидуальных особенностей данного лица.'Далее, индивидуальность сказывается в более или менее неодинаковом влиянии тех или других из вышеуказанных условий опыта, например, предшествующего кратковременного и длящегося света, а также неодинакового влияния неравномерных антрактов между раздражениями и т. п.
ИССЛЕДОВАНИЕ ФУНКЦИЙ МОЗГОВОЙ КОРЫ
С ПОМОЩЬЮ ЕСТЕСТВЕННЫХ СОЧЕТАТЕЛЬНЫХ
РЕФЛЕКСОВ. ЗНАЧЕНИЕ ЭТОГО МЕТОДА
ПО ОТНОШЕНИЮ К ЦЕНТРАМ ВНУТРЕННИХ ОРГАНОВ
И РАЗЛИЧНЫХ СЕКРЕЦИЙ
Кроме обычных расстройств двигательной сферы, служащих выражением паралича движения и чувствительности, очень важными указателями нарушения функций мозговой коры являются,, с одной стороны, обыкновенные рефлексы, с другой —специальные сочетательные рефлексы.
Значение последних особенно важно при исследовании корковых центров для движения растительных органов и корковых секреторных центров, которые за последнее время были предметом исследования по методу раздражения преимущественно в нашей лаборатории. Ввиду этого мы остановимся здесь подробнее на применении метода сочетательных реакций к изучению упомянутых центров мозговой коры.
Специальные сочетательные рефлексы состоят во внешней реакции, обусловленной не силой и не качеством внешнего раздражения, а установившимся соотношением его с другим деятельным, т. е. возбуждающим известный эффект, раздражением,-благодаря бывшим ранее комбинациям этих раздражений. Так, звук бича возбуждает резкую двигательную реакцию не сам по-себе, так как другие подобной же силы и даже сходственные по характеру звуки не возбуждают аналогичной реакции, а как звук, сочетанный с бывшим ранее ударом по спине, произведенным таким же бичом или аналогичным орудием; равным образом шипение змеи возбуждает в нас известную двигательную реакцику не само по себе, а как звук, сочетанный с возможным укусом, так как всякое другое шипение ничуть не возбуждает подобной: же реакции. Вид ружья заставляет нас отвернуться от дула, так как дуло ружья, само по себе не содержащее ничего' страшного, сочетается с возможным выстрелом и следовательно, с известной опасностью. Звук тарелок возбуждает обильное-слюноотделение во время голода опять-таки не самим звуком^ а тем, что этот звук связан с бывшей едой. Звук того или другого слова возбуждает реакцию не силой звука, а благодаря сочета-
16?»
нию его с другими внешними раздражениями. Вид кошки возбуждает в собаке сильную двигательную реакцию опять-таки вследствие бывшего ранее действия когтей кошки на спине или морде собаки. Этот род рефлексов мы называем сочетательными рефлексами, именно благодаря тому, что они являются результатом сочетательной деятельности мозговой коры.
В лаборатории И. П. Павлова эти явления в области слюноотделения называют «естественными условными рефлексами»; но я не вижу никакого основания держаться этого термина, так как понятие «условности» в данном случае мало что-либо говорит в пользу характеристики этих рефлексов, —тем более, что связь эффекта с раздражением здесь вовсе не «рыхлая», как полагается это принимать по отношению к так называемым «искусственным условным рефлексам», а напротив того, эти рефлексы, развиваясь под влиянием бывших ранее сочетанных раздражений, в известных случаях могут отличаться поразительным постоянством.
Возьмем ту же реакцию на бич. Достаточно хотя однажды собаке почувствовать влияние бича на своей спине, чтобы вызванная им реакция затем повторялась многое число раз с поразительным постоянством.
В упомянутых сочетательных рефлексах, как показывают исследования, несомненно, принимает участие мозговая кора.
Если мы удалим у собаки височные доли с обеих сторон в области так называемого слухового центра, то оказывается, что животное становится безразличным к звукам бича, зову и пр., тогда как оно еще реагирует на звуки общей двигательной реакции (исследования Ферье, Мунка, Лючиани, мои и др.).
Точно так же, если у животного удалить обе затылочные доли, то хотя оно и может обходить препятствия, но оно не реагирует на поставленную перед ним пищу, а равно и не реагирует насвоего хозяина (исследования Ферье, Мунка, Лючиани, мои и других авторов).
В этом смысле говорят и многочисленные клинические исследования с так называемой «психической» глухотой и «психической» слепотой.
Должно, однако, иметь в виду, что так дело представляется лишь при удалении так называемых чувственных, resp. воспринимающих, центров мозговой коры, но ничуть не представляется возможным переносить полностью эти результаты и на двигательную сферу мозговой коры, расположенную на сигмовидных, resp. центральных, извилинах.
Дело в том, что двустороннее удаление двигательных областей не вполне устраняет осуществление этой реакции, так как животное в этом случае реагирует еще на звук бича и на вид пищи или своего хозяина, но так как двигательная его сфера утратила известную часть мимики и лишилось отдельных движений, то и не обнаруживается на звук бича в этом случае всей той полноты двигательной реакции, которая наблюдалась до операции.
170
Так, животное с удаленными двигательными областями при звуках бича убегает от него, а при виде хозяина подходит к нему, но не выражает испуга в первом случае, ни ласки во втором случае. Если бросить собаке кость, то она тотчас же набрасывается на нее, но не может пользоваться своими лапами, чтобы удерживать кость между ними.
Иначе говоря, в двигательной реакции на внешние раздражения животное испытало соответственный ущерб, выражающийся утратой части своей мимики и утратой отдельных движений, но реакция в более элементарном виде все еще проявляется. Факты эти объясняются, очевидно тем, что для движения мы имеем центры не в одной только сигмовидной извилине, но в других частях полушарий, так как удаление сигмовидной извилины не лишает животного ходьбы и бега.
Исследование сочетательных рефлексов приобретает особенную важность, между прочим, по отношению к изучению корковых центров, имеющих отношение к иннервации внутренних органов, а равно и по отношению к секреторным центрам мозговой коры.
Дело в том, что растительные и в том числе секреторные функции имеют назначение выполнять наиболее важные жизненные потребности организма, обеспеченные простыми рефлексами. Благодаря этому, удаление соответствующих корковых центров не отражается существенным образом на проявлении относящихся к ним элементарных функций организма, а между тем известный недочет в более сложных проявлениях нервной деятельности в отношении этих функций выступит тотчас же, как только мы обратимся к исследованию сочетательных рефлексов.
Возьмем дыхательные центры. Ныне не может подлежать сомнению, что, кроме спинномозговых и основного координирующего дыхательного центра, заложенного в продолговатом мозгу и управляющего дыханием при посредстве спинномозговых центров, имеются подкорковые центры, заложенные в области среднего и межуточного мозга, и, кроме того, имеются еще и корковые центры дыхания, указываемые рядом авторов и подробно исследованные у нас доктором Жуковским1.
Роль спинномозговых и основного координирующего центра продолговатого мозга в настоящее время представляется уже достаточно выясненной.
Имеются факты, говорящие в пользу важной рефлекторной деятельности и подкорковых дыхательных центров среднего и межуточного мозга 2.
Но роль корковых центров дыхания представлялась до позднейшего времени неясной, тем более, что вслед за их удалением не обнаруживается видимых расстройств в деятельности
1 Жуковский, Диссертация, СПБ, 1898.
2 В. Бехтерев, Основы учения о функциях мозга, в V.
171
дыхания. Однако исследования не оставляют сомнения в том, что корковые дыхательные центры играют известную роль в отношении сочетательных рефлексов.
Так, производившиеся у нас опыты показали, что вслед за удалением дыхательных корковых центров в обоих полушариях не только сохраняются общие рефлексы на дыхание, получающиеся, например, раздражением п. ischiadici и твердой оболочки, но и раздражение таких нервов, как обонятельного, электричеством и специфическими раздражителями (сероуглерод, жженое перо и пр.) вызывают еще экспираторную задержку дыхания. Равным образом и раздражение слухового и зрительного нервов вызывало инспирацию с замедлением дыхательного ритма.
Далее исследования показали, что в вышеупомянутом случае удаления у животного дыхательных корковых центров в обоих полушариях приближение кошки к собаке уже не производит того обычного эффекта на дыхание, который наблюдается у здоровых животных (Жуковский) 1.
Ясно, что мы имеем здесь дело с центрами, играющими известную роль в отношении так называемых психорефлекторных, точнее —сочетательно-рефлекторных движений со стороны дыхания.
Играют ли эти центры исключительную роль в деле передачи этих рефлексов или же, что вероятнее, возможно со временем образование для этих рефлексов других путей, которые постепенно компенсируют вышеуказанный недочет, —это нуждается еще в дальнейшем выяснении.
Не менее поучительные факты известны и по отношению к центрам половых органов, функция которых теснейшим образом связана с состоянием сосудистой системы.
Исследования, произведенные у нас (мною и доктором, Л. М. Пуссепом2), не оставляют сомнения в том, что в коре полушарий в пределах так называемой двигательной области имеется особый центр половых органов, существующий для обоих полов. У кобелей этот центр помещается в верхней трети gyr. posteruciati вблизи центра хвоста.
Роль полового центра опять-таки выясняется при испытании сочетательного полового рефлекса. Дело в том, что и после двустороннего удаления вышеуказанного центра, как показали у нас опыты доктора Пуссепа, половая возбудимость не исчезает, так как при механических раздражениях в области полового органа последний обнаруживает нормальную и даже повышенную возбудимость; но тем не менее естественные возбудители полового чувства в период течки у собак, как они даются в природе зри-
1 Жуковский, Диссертация, СПБ, 1908.
2 Бехтерев В., Основы учения о функциях мозг,), в. VI..
172
тельными раздражениями, оказываются уже недействительными, и кобель кажется при этих условиях, по крайней мере, вначале, утратившим свою половую возбудимость (Пуссеп) х-
Имеются положительные указания в пользу того, что и зрачковые центры, заложенные в передней части полушарий, играют известную роль при выполнении естественных сочетательных рефлексов. Известно, что зрачки расширяются при внешних раздражениях, приводящих животное в испуг. При этом оказывается, что предварительная перерезка п. oculomotorii не устраняет расширяющего зрачок влияния раздражений, возбуждающих аффект, откуда очевидно, что здесь дело идет об активном расширении зрачка, а не обусловленном одним только угнетающим действием на центры, сокращающие зрачки.
Если мы затем удалим у животного корковые центры зрачков в лобной области, то оказывается, что определенные внешние раздражения, приводящие животное в испуг, уже не вызывают расширения зрачков (Браунштейн)2.
При этом случае необходимо, однако, заметить, что вопрос ■о зрачковых центрах в действительности не так прост, как это может показаться с самого начала, так как, кроме лобных зрачковых центров, имеются еще затылочные и теменные зрачковые центры 3, которые, без сомнения, также должны играть известную роль в отношении сочетательных зрачковых рефлексов, но какие именно рефлексы передаются при посредстве тех и других центров—этот вопрос мы должны предоставить еще будущим исследованиям.
Далее, как уже ранее упоминалось, метод специальных сочетательных рефлексов имеет применение и к исследованию секреторных центров мозговой коры.
По крайней мере, по отношению к нескольким исследованным у нас секреторным центрам удалось с успехом применить метод исследования специальных сочетательных рефлексов. Я говорю о слюноотделительном, желудочно-сокоотделительном и молочноотделительном центрах, роль которых в этом отношении была указана исследованиями, производившимися в нашей лаборатории (Бари4, Спиртов5, Белицкийв, Гервер7, Никитин 8).
1 Бехтерев В., Основы учения о функциях мозга, 1906, стр. 1055—1093.
2Браунштейн, К учению об иннервации движений зрачка. Диссертация, 1893.
3 Бехтерев В., Основы учения о функциях мозга, в. VII.
4 Бари, Неврологический вестник, 1899, VII. 'Спиртов, Научное собрание врачей клиники душевных болезней, май 1908.
6 Белицкий, Обозрение психиатрии, 1905.
7 Гервер, Обозрение психиатрии, 1906.
8 Никитин, Научные собрания врачей клиники душевных болезней, 1906; его же диссертация, СПБ.
173
Допустим, что мы удалили у собаки с выведенным наружу протоком слюнных желез (подчелюстной и parotis) установленный нами1 и затем более точно локализированный работой доктора Спиртова корковый слюноотделительный центр с обеих сторон2. Испытывая оперированное животное на обычные рефлексы, например, путем дачи ему еды, мы убеждаемся, что слюна течет из трубок каждый раз, когда животному дают что-нибудь съесть. Равным образом, если мы будем вливать животному слабый раствор соляной кислоты, слюна потечет из трубок каплями. Точно так же и обонятельные раздражения способны вызвать отделение слюны.
Но в отличие от нормальных животных, у оперированных вышеуказанным образом собак в ближайшие после операции дни, когда животное сделается доступным для исследования, обнаруживается следующий факт. Если мы будем показывать им на расстоянии непахучие пищевые продукты (сахар) или хотя бы и пахучие (мясо), но плотно закупоренные в банке, то животное, как бы голодно оно ни было, не обнаруживает слюноотделения. Равным образом, стук тарелки или склянки, из которой давалась пища оперированному животному, не будет вызыввть у него слюноотделения, несмотря на то, что в здоровом состоянии те и другие раздражения у животного в периоде голода возбуждали обильное слюноотделение, особенно после того, как вместе с показыванием пищи звуки посуды повторялись неоднократно при еде животного 3.
Только что указанный эффект наблюдается в течение первых 5—6 дней, после чего наступает постепенное восстановление нормальных отношений (доктор Спиртов).
Таким образом, здесь дело идет о временной утрате тех же сочетательных рефлексов, которые, как мы- видели выше, выпадают и при удалении корковых центров для движения внутренних органов.
Вышеуказанные данные относительно слюноотделительного центра оспаривались доктором Тихомировым при исследованиях с так называемыми искусственными условными, resp. сочетательными, рефлексами4,'но без достаточных оснований, так как и по описанию, и по рисункам, приложенным к работе, можно видеть, что автором не удалялся весь слюноотделител!--
1 В одном из майских заседаний научных собраний клиники душевных болезней за текущий год при большом собрании врачей и студентов была произведена демонстрация по методу раздражения упомянутого центра доктором Спиртовым (из нашей лаборатории), при исследованиях которого выяснилось, что наиболее деятельной частью коры в отмеченной нами области является gyr. сотр. anter.
2 Бехтерев иМиславский, Основы учения о функциях мозга, в. VI.
3 Доктор Белицкий.
4 Тихомиров, Опыт строго объективного исследования мозговых функций. Диссертация, СПБ.
174
ный корковый центр, а оставлялась неудаленной вполне самая важная его часть, именно: gyr. сотр. ant.
В устранение вышеуказанного возражения я предложил доктору Спиртову вновь проделать исследование над выяснением роли корковых слюноотделительных центров, причем в результате этого исследования факты, сообщаемые ранее из нашей лаборатории, вполне подтвердились.
При этих исследованиях прежде всего выяснилось, что из всей области, отмеченной ранее нами в виде слюноотделительной, чаще всего особенно деятельной в смысле вызывания слюноотделения является область gyri compositi ant. Если эту область устранить, причем, очевидно, дело идет об устранении ничуть не всей слюноотделительной области, а лишь ее наиболее деятельной части, то в течение вышеуказанного периода у животного не удается вызвать ранее хорошо получавшихся естественных слюноотделительных рефлексов на звуковые и на зрительные раздражения, тогда как на обоняние, а тем более на вкус они получаются хорошо.i
Предположение, что дело сводится здесь к одному угнетению подкоркового слюноотделительного центра, опровергается следующим образом: специальные исследования на раствор кислоты, возбуждающий подкорковый слюноотделительный рефлекс, показали, что у животного раствор кислоты в течение всего послеоперационного периода возбуждает слюноотделение, причем он должен быть вслед за операцией несколько усилен, но это продолжается лишь 2—3 дня в период общего угнетения собаки, после чего вполне восстановляется прежняя, т. е. бывшая до операции, возбудимость подкоркового слюноотделительного рефлекса, тогда как сочетательный корковый рефлекс на зрение и слух остается еще утраченным в течение, как упомянуто, 5—6 дней.
Должно иметь в виду, что вышеуказанный эффект относится только к естественным рефлексам, о чем и заявлялось ранее в работе доктора Белицкого; между тем как путем многократного вливания животному в рот кислоты и возбуждения этим путем подкоркового слюноотделительного центра одновременно со зрительным раздражением пищей удается через некоторое время снова привить животному сочетательный зрительный слюноотделительный рефлекс. Этот факт может быть объяснен тем, что и помимо корковых секреторных центров, имеющих в указанном отношении несомненное значение, упомянутый рефлекс при последовательном упражнении животного находит иные пути для своего обнаружения, очевидно передаваясь в этом случае непосредственно от зрительного и слухового коркового центра к подкорковому слюноотделительному центру.
Точно так же и по отношению к корковому центру желудочного сокоотделения, легко демонстрируемому по методу раз-
175
дражения, работой доктора Гервера (из нашей лаборатории) "было доказано, что вместе с удалением этого центра у животного прекращается отделение так называемого психического сока (по терминологии И. П. Павлова), которое обнаруживается при раздражении животными пищевыми продуктами на расстоянии, тогда как другие рефлексы на сокоотделение при этом остаются сохраненными.
И этот факт был оспариваем из лаборатории И. П. Павлова, но работа, производимая ныне у нас доктором Трекером со специальною целью еще раз проверить полученные результаты, вполне подтвердила ранее сообщенные данные, причем выяснилось, что отсутствие сочетательного сокоотделительного рефлекса на показывание пищи у оперированных собак наблюдается лишь в течение первых нескольких дней, приблизительно около 6—8, тогда как у контрольных животных, которым производилось удаление других, хотя бы и соседних частей мозговой коры, ничего подобного не наблюдалось. Но и с течением времени еще обнаруживается у некоторых животных, по наблюдениям доктора Трекера, заметная количественная разница между собакой с удалением коркового желудочного центра и собакой, оперированной в других областях коры, в смысле более позднего появления желудочного сока при раздражении пищей на расстоянии у первой собаки. Заслуживает внимания, что собаки с удаленным сокоотделительным центром в первое время вообще менее охотно едят по сравнению с оперированными в других частях мозговой коры и при показывании им пищи даже отворачиваются от нее.
Тот факт, что с течением времени эффект операции постепенно выравнивается, служит объяснением тому, почему другие авторы не наблюдали этих явлений при удалении коркового сокоотделительного центра. Дело в том, что, по заявлению И. П. Павлова в майском научном собрании врачей клиники душевных болезней, исследование сочетательного сокоотделительного рефлекса в его лаборатории начиналось лишь с 6-го дня, так как до этих пор оперированные собаки не принимали пищи вовсе.
Далее работой доктора Никитина из нашей лаборатории было доказано по методу раздражения в ряде опытов над молочными некураризированными и кураризированными овцами существование особого молочноотделителыюго центра в передних отделах мозговой коры, значение которого также было выяснено с помощью исследования сочетательных рефлексов. Оказалось, что вместе с удалением этих центров в обоих полушариях мозга у молочной овцы в первое время не выделяется молоко ни при виде сосущего ее ягненка, ни при обнаруживании им своего голоса, что наблюдалось с постоянством у того же животного до произведенной операции. Для выяснения того,
176
как долго длится этот эффект, должны быть поставлены еще особые опыты.
Все вышеизложенное не оставляет сомнения в том, что для исследования функций корковых центров метод испытания специальных сочетательных рефлексов получает особое значение: благодаря ему, удается выяснить физиологическое значение целого ряда корковых центров и в том числе секреторных, роль которых иначе оставалась бы совершенно неясною, так как с удалением их в более элементарных отправлениях нервной системы не происходит какого-либо нарушения в отправлениях организма.
12 В. М. Бехтерев
Ill
ТРУДЫ
по
НЕВРОПАТОЛОГИИ
12«
НАБЛЮДЕНИЯ ТРОФИЧЕСКИХ ИЗМЕНЕНИЙ КОЖИ В ЗАВИСИМОСТИ ОТ РАССТРОЙСТВ НЕРВНОЙ СИСТЕМЫ1
1. Сыпи, распространяющиеся в области пораженных нервов
В смысле расширения наших знаний относительно трофического влияния нервной системы наиболее убедительными представляются те случаи поражения кожных покровов, которые ограничиваются определенными нервными участками. В особенности интересны в этом отношении некоторые формы сыпей. Так, с давних пор известно, что одна из пузырчатых сыпей —herpes zoster —распространяется весьма часто по ходу пораженных нервов. Впервые связь этой сыпи с поражением нервов была указана Парро в 1857 г.; вслед за тем то же самое было подтверждено прекрасными наблюдениями Руже, Шарко, Готара, Эсмарха, Береншпрунга и многих других. Действительно, указанное распространение этой сыпи настолько постоянно, что в настоящее время считается одним из наиболее характерных ее признаков. Почти всегда она ограничивается исключительно одной половиной тела, не переходя срединной линии как спереди, так и сзади. Далее, какая бы область тела ни была занята этой сыпью, она всегда следует довольно точно ходу нервных ветвей: так, на груди она распространяется по направлению межреберных нервов, и если занимает второе и третье пространство, то очень часто вместе с тем идет и по внутренней поверхности плеча, соответственно анастомозу, посылаемому вторым и третьим межреберными нервами в подкожную плечевую область. На шее сыпь идет по всем направлениям, соответственно веточкам plexus cervicalis, на лице —соответственно ветвям п. trigemini; на руках и на ногах —длинными рядами в направлении подкожных нервов. Кроме того, относительно этой сыпи известно, что она составляет весьма частый спутник невралгий, в особенности у взрослых; появление ее в этих случаях часто совпадает по времени с наступле-
1 Сообщение, сделанное на заседании Психиатрического общества 16 мая текущего года (с демонстрацией больных).
181
нием невралгического приступа, иногда незадолго предшествует или следует за ним. Но herpes zoster, несмотря на частоту ее появления при различных нервных поражениях, далеко не составляет единственную сыпь, распространяющуюся по ходу пораженных нервов. Положительные наблюдения указывают, что иногда подобное же распространение принадлежит сыпи, отличающейся всеми характерными признаками pemphigus. Сюда принадлежит известный случай Шарко \ Также Деже-рин 2 при симметричном склерозе боковых пучков наблюдал высыпание пузырьков этой сыпи в области sacrum, затем на обеих верхних и потом на нижних конечностях. При вскрытии этого субъекта, погибшего от маразма, были найдены патологические изменения в нервах конечностей на уровне развития пузырьков. Затем Бриссо сообщил наблюдение о гемиплегике, подверженном начинающейся атрофии мускулов плеча, у которого наступило высыпание пемфигоидных пузырьков на парализованной половине туловища; при этом конечности оставались не пораженными, никаких болей у больного не существовало. Положение и распространение сыпи в данном случае •было настолько характерно, что автор не без основания предложил дать этой редкой форме название «Pemphigus zoster». К той же категории фактов относятся наблюдения относительно других форм накожных сыпей и, между прочим, крапивной сыпи. Шарко по этому случаю говорит: «Нередко случается видеть, что кожа голени и бедра временно покрывается ■более или менее сливною папулезною и лишаевидною сыпью вслед за приступами стреляющих болей, характеризующих атаксию (ataxie locomotrice). У одной женщины, находящейся на излечении в больнице Сальпетриер, каждый раз после приступа появляются большие бляшки крапивной сыпи на уровне тех мест, в которых ощущалась самая сильная боль» 3. Также Вуль-пиан 4 у своих больных наблюдал высыпание розеолезнои сыпи на туловище, совпадающее с наступлением crises gastriques. •Случаи, аналогичные предыдущим, известны и в истерии δ. При травматических же повреждениях периферических нервов эритематозные сыпи составляют явление далеко не редкое. Наконец, в тех же случаях весьма часто появляется страдание кожи, известное у английских писателей под названием glossy skin, выражающееся утончением кожи, особенною гладкостью и блеском эпидермиса. В других случаях, напротив, наблюдают чрезмерное утолщение с опуханием кожи и подкожного
1 Charcot, Journal de physiologie, 5 Janvier 1859, t. II; Болезни нервной системы, т. I, стр. 19.
2 D е j ё г i η e, Paralysie generale, troubles trophiques cutanes. Pemphigus, Arch, de physiologie, 1876, p. 307.
3 Шарко, Болезни нервной системы, т. I, стр. 66.
4 V и 1 р i a η, Maladies de la moelle, Observ. d'ataxaie locomotrice 6 Annales de dermatologie, 1880, 20.
182
слоя, нередко сопровождающееся более или менее значительным отеком.
Относительно экземы мы еще не имеем положительных данных с указаниями на связь ее с нервными поражениями. Впрочем, одно наблюдение, занесенное в литературу, заслуживает быть упомянутым. «Человек 60 лет никогда не страдал экземой. Сильный ушиб плеч. Боли, распространяющиеся по длине плеча; 40 часов спустя "появление, экземы по направлению п. га-dialis, быстро сделавшейся сливною и прошедшей в течение 15 дней без последовательного рецидива» х.
Зато несомненные сведения в том же направлении имеются относительно ichtyosis и ectimae. Уже американские хирурги наблюдали, что вследствие травматических поражений нервов кожа может принять вид настоящего ихтиоза. На основании некоторых данных Арнозан предполагает, что во всех подобных случаях ichtyosis соответствует развитию медленно идущего неврита 2. Лелуар 3 недавно сообщил также о двух случаях местного ихтиоза вследствие неполных ранений нервов. Мы не приводим здесь других аналогичных фактов, которых в литературе имеется множество. Также всем более или менее известны случаи появления ихтиоза у старых параплегиков. Но вот наблюдение, принадлежащее Дюге, которое заслуживает в этом отношении особенного внимания. Больной с детства страдал ichtyosis serpentina, представлявшей характерное расположение в форме лент параллельно межреберным пространствам. Поражение не переходило срединной линии ни спереди, ни сзади и занимало собою семь или восемь межреберных промежутков, оставляя между ними-параллельные участки здоровой кожи 4.
Что касается эктимы, то уже давно известно, что у табетиков она иногда распространяется соответственно различным нервам по коже нижних конечностей. Шарко передает следующий факт: женщина 61 года 8 лет тому назад была принята в больницу Сальпетриер с атрофией оптических нервов; в настоящее время она одержима чрезвычайно характеристической двигательной атаксиею. Болезнь развивалась очень быстро. Один из приступов болей, случившийся первого июня 1867 г. был особенно сильный. Жестокие боли в течение нескольких дней были ощущаемы больною на"пути кожных ветвей малого седалищного нерва и правого нерва m. levateris ani. В то же
1 Brouardel, cit. par Arnozan, De lesions trophiques consec. aux maladies du systeme nerveux, These, Paris, 1880, p. 124.
2 См. случаи Arnozan и Eulenburg в Deutsche klini-sche Wochenschrift, 1871; A r η о z a η, [Des lesions trophique, These, 1880, 151.
3 L e 1 о i r, Arch, de physiol. norm, et pathol., 1880, № 3, pp. 409 et 411.
4 Duguet, cit. par L e 1 о i r, Arch, de physiol. norm, et pathol., 1881, № 3, p. 411.
183
время соответствующие места кожи покрылись многочисленными, аналогичными с эктимой, пустулами, из коих некоторые превратились в глубокие изъязвления. Кроме того, в крестцовой области с правой стороны, на расстоянии нескольких сантиметров от срединной линии и непосредственно над оконечностью копчика, образовалась кругловатая эшара, имевшая около 5 см в диаметре и занимавшая почти всю толщу кожи. Эта язва по отделении омертвевших частей зажила только в конце второго месяца. В другой раз приступ стреляющих болей распространился по направлению вертикальной ветви п. sapheni interni sinistri и вскоре на этом пространстве появилась пустулезная сыпь» *.
Известны также другие примеры кожных поражений, находящихся в прямой зависимости от страданий нервной системы.
Что касается теории и патологоанатомической основы страдания во всех подобных случаях, то сведения наши в этом отношении крайне ограниченны. Тем не менее, для того чтобы интересующиеся могли лучше ориентироваться в этом предмете, считаю нужным привести здесь взгляды некоторых авторов и результаты вскрытий, которые до сих пор известны. Из того факта, что часть указанных изменений в коже нередко сопровождается сильными чувствительными раздражениями (как, например, стреляющими, молниеносными болями при tabes), выводили заключение, что причина их находится в раздражении трофических волокон. Такой взгляд по преимуществу был развиваем Шарко и его учениками. В пользу этого мнения приводили в особенности частое наступление трофических расстройств в коже при травматических повреждениях, неполных перерезках и ушибах нервов, вообще при всех условиях, производящих ирритативный процесс в нерве или воспаление, и указывали на появление herpes при невралгиях и neuritis. Но, с другой стороны, существуют несомненные как патологические, так и физиолого-экспериментальные факты, доказывающие, что аналогичные изменения в коже могут происходить и при условиях, в которых становится в высшей степени вероятным существование паралича трофических нервов. Таким образом, по всей вероятности, как функциональное возбуждение, так и инертность трофических волокон могут приводить к ну-тритивным изменениям в коже, хотя до сих пор мы не имеем возможности определить, в частности, какое участие при каждом из этих изменений принимает то или другое явление. Относительно анатомической стороны вопроса известные до сих пор факты приводят к следующим выводам. Периферические пути для трофических импульсов, идущих к коже, без сомнения, лежат в чувствительных волокнах. В пользу этого говорят с по-
1 Шарко, Болезни нервной системы, т. I, стр. 65 и 66.
184
ложительностью результаты многих вскрытий. При herpes zoster, например, нередко находили сдавливание нерва опухолями, как в случаях Гернера и Шиффера, абсцессом в случае. Эсмарха, раком в случае Оливье. Далее, Броун-Секар указал на менингит, который мог действовать раздражающим образом на задние корешки \ Наконец, известно множество случаев травматического миэлита, сопровождающегося появлением herpes на местах кожи, соответствующих распространению пораженного нерва 2. Вопрос наиболее важный и в то же время наиболее трудный, где помещаются трофические центры кОжи, в настоящее время еще не может быть решен утвердительно. Здесь достаточно только сказать, что французская школа, основываясь на результатах некоторых вскрытий, обращает внимание, с одной стороны, на серые задние столбы спинного мозга, на так называемые внутренние лучистые пучки (fais-ceaux radiculaires internes) 3, с другой стороны ■—на спинномозговые ганглии 4.
Наблюдение, которое недавно мне удалось сделать, благодаря особенной любезности проф. Полотебнова, относится к случаю с несомненным трофическим поражением кожи в виде сыпи, распространяющейся по направлению нервов нижних конечностей. Случай этот, как мне кажется, не лишен интереса и заслуживает быть приведенным 'наряду со всеми вышеупомянутыми.
Больная П., женщина 50 лет, пришла на амбулаторный прием в клинику 7 мая. Мать ее умерла в холере, отец — от апоплексического удара; две сестры всегда были нервные; из них старшая умерла в чахотке. В детстве,будучи лет 10, больная перенесла какую-то общую болезнь с опуханием всего тела. Кроме того, она всегда страдала частыми головными болями и расстройством желудка, что продолжается и до сих пор. Наконец, с давних пор больная имеет припадки головокружения, сопровождавшиеся тошнотой и даже рвотой. В последние годы она начала жаловаться
1 В а г t h, Etudes de l'herpes, Mernoire pres. au concours pour les prix de Pinternat, 1878.
г Относительно pemphigus ср. цитированный случай Дежерина.
3 Ш а р к о, Болезни нервной системы, т. I, стр. 65.
4 Arnozan, De lesions trophiques consec. aux maladies du systeme nerveux, These, Paris, 1880, p. 130 e. suiv. В литературе имеется уже несколько счастливых вскрытий, в которых herpes zoster можно было поставить в связь с поражением спинномозговых ганглиев. Таковы случаи Шарко (Болезни нервной системы, т. I, стр. 69, 70), Висса (Arch. d. Heil-kunde, 1871, Bd. IV und V), Затлера (Vierteljahrschrift f. d. Mat. und Syph., 1878), Шанделе (Arch, de physiol. norm, et pathol., 1879, p. 674) и др.
В последнее время Лелуар (Contribution а 1 'etude des affections cuta-nees d'origine trophique, Arch, de physiol. norm, et pathol., 1881, № 3, p. 391), задавшись целью определить причинное нервное поражение, лежащее в основе трофических кожных изменений, находил в случаях ichtyosis и ectimae дегенеративные явления в кожных нервах и отчасти в задних корешках (последние, по предположению автора, может быть, составляют последствие поражения межпозвоночных ганглиев, которые, к сожалению, не были исследованы).
J 85
на частые судороги в нижних конечностях, в особенности в икрах, что иногда случается и теперь, но в слабой степени. Года четыре или пять тому назад появилась боль в пояснице, сопровождаемая чувством сдавли-• вания; боль особенно беспокоит больную при усидчивом труде. В последнее время, кроме того, довольно часто появляются боли в области живота, а с неделю тому назад боль распространилась на правую половину таза с наружной его стороны, продолжаясь в течение 4 дней кряду. Иногда невралгические боли появляются и в конечностях. Так, 3 дня тому назад больная испытывала сильную боль в стопе правой конечности. Прежде, до наступления этих болей нередко появлялись различного рода парестезии в виде ползающих мурашек в различных частях тела, преимущественно в спине и в конечностях. Год тому назад во время развития сильных опоясывающих болей в области живота наступило продолжительное задержание мочи, причем в течение около двух месяцев моча выходила каплями. Кроме того, с давних пор больная страдает сильными запорами. В настоящее время, кроме всех указанных симптомов, у больной иногда появляются приступы сердцебиения, случавшиеся и прежде. Головокружения в настоящее время почти вовсе не появляются, но иногда наступает тошнота. Чувство общей тягости постоянное. Больная чувствует слабость в конечностях и при ходьбе быстро утомляется. Сон всегда тревожный, и больная просыпается почти каждый час, чувствуя при этом общее недомогание и боли в руках, спине и ногах. Аппетит часто расстраивается. Регул больная не имеет уже два года. В последнее время у нее появилась сыпь на всей внутренней стороне правой ноги, сопровождаемая зудом; от расчесов, впрочем, больная строго воздерживалась.
П. женщина среднего роста, худощавая; слизистые оболочки губ и век довольно бледные, кожа повсюду сухая и вялая. На лице и в особенности на шее замечается легкая■ темножелтоватая пигментация кожи которая, по словам больной, наступила в последние два года. Такого же рода пигментация замечается на плечах рук, на туловище и на ногах, но в меньшей степени. На правой стороне лба небольшой шрам вследствие сильного ушиба о мебель, случившийся года два тому назад. В волосах замечается незначительная седина. Небо слегка седлообразное, верхние резцы испорчены; язык покрыт маленькими язвочками. В легких прослушиваются хрипы и местами жесткое дыхание: в сердце ничего патологического. Печень выстоит на полтора пальца из-под края ребер по мамми-лярной линии и болезненна. Правая почка подвижна и прощупывается; в моче отсутствие белка и сахара. На пояснице, в области glutaei в особенности же в окружности anus и на местах, соответствующих гребешкам подвздошных костей, обращают на себя внимание густо рассеянные пятна темнобурого цвета в виде пигментированных рубцов; такого же рода пятна, местами сливающиеся, распространяются в виде полосы вдоль всей внутренней поверхности правой ноги. Начинаясь от паховой области, эти пигментированные пятна полосой спускаются вниз по внутренней поверхности бедра; затем, обходя с задне-внутренней стороны колена, переходят на задне-внутреннюю поверхность голени, оканчиваясь вершка на 3 выше malleolus internus; в виде продолжения этой полосы несколько рассеянных пятен замечается также на внутренней и верхней поверхности стопы. Точное исследование топографии этой полосы пигментированных Рубцовых пятен показало, что она соответствует распространению п. ileo-hypogastrici и кожных ветвей п. sapheni majoris, преимущественно п. cutaneus femoris interni и п. surae interni. Кроме столь характеристично расположенных пигментированных рубцовых пятен, в небольшом количестве встречаются подобного же рода пятна, рассеянными иа других частях правой, также левой нижних конечностях, в особенности ниже колен, на стопе и, наконец, в локтевых сгибах обеих верхних конечностей. Относительно характера этих пигментированных пятен я должен сказать следующее: больная в первый раз в начале апреля явилась на амбулаторный прием в клинику проф. Полотебнова с пигментированными пятнами на правой ноге в виде той же полосы, как мы ее видим теперь. В пояс-
186
яичной области, между тем, этих пятен не замечалось, а вместо них была замечена сыпь в виде плоских пупырышек со всеми характерными признаками lichen ruber и была диагносцирована за таковую профессором Поло-тебновым. Больной был назначен мышьяк в виде пилюль. Затем, в следующий раз, несколько недель спустя на месте указанной сыпи в поясничной области обнаружились такого же рода пигментные рубцовые, слегка углубленные пятна, как и на ноге. Следовательно, с большей вероятностью можно было заключить, что пигментированная полоса, начинающаяся в области кожных ветвей п. ileo-hypogastrici и спускающаяся вниз по направлению ветвей п. sapheni majoris, составляет след протекшего lichen ruber. Впоследствии вновь появившиеся незначительные высыпания по соседству с Рубцовыми пятнами вполне подтвердили точность сделанного заключения. Из явлений со стороны нервной системы, кроме данных, указанных в анамнезе, замечается еще следующее. На каждой ноге в отдельности при закрытых глазах больная удерживается заметно хуже, нежели при открытых. Также походка с закрытыми глазами немного шаткая и нетвердая. Сила обеих рук одинакова. В общей чувствительности кожи резких изменений не замечается. Сухожильные рефлексы с ligament, patellare заметно усилены. Спинные позвонки в поясничной области при давлении немного болезненны. Также получается довольно резкая боль в области, соседней с crista ilei, при давлении по направлению нервов поясничного сплетения и по всей правой ноге вдоль п. ischiadicus и в особенности cruralis.
Итак, у нашей больной, судя по анамнезу и данным исследования, можно с положительностью заключить о центральном спинномозговом страдании. Вместе с тем у ней замечаются пигментированные рубцы от бывшей lichen ruber в поясничной области и по всей вероятности такого же происхождения рубцы, распространенные преимущественно в направлении ветвей болезненного при давлении п. cruralis. Такое характеристическое распределение Рубцовых пятен как последствия протекшей сыпи по болезненным нервам правой нижней конечности доказывает с несомненностью их трофическую природу и совершенно аналогично вышеприведенным случаям с другими формами сыпей. Что касается характера самой сыпи, то, насколько мне известно, в литературе еще нет никаких указаний относительно распространения lichen ruber по направлению кожных нервов. Эта форма сыпи, впервые описанная Гебра, как болезнь sui generis еще до сих пор остается весьма мало изученною, в особенности со стороны ее этиологии. Мы знаем, что lichen ruber не обусловливается конституциональными моментами, так как поражает видимо здоровых людей. Кроме того, известно, что она не заразительна и не наследственна х. На основании же приведенного наблюдения возможно предполагать, что по крайней мере для некоторых случаев этой болезни, расстройство нервной системы играет видную роль в этиологии.
1 К а п о з и, Патология и терапия накожных болезней, перевод Кузнецова, Харьков, 1881, стр. 325.
187
II. А н о м а л и я кожного пигмента
Количественное изменение кожного пигмента в некоторых случаях поражений нервной системы обращало на себя внимание многих авторов. Наиболее замечательным примером в этом отношении может служить случай Ростана, приводимый Ли-коком х: женщина, приговоренная к смерти во время французской революции, быстро почернела под влиянием сильного нервного потрясения: окраска кожи в этом случае оставалась до самой ее смерти, наступившей 30 лет спустя после отмены смертного приговора. Барлоу2в доказательство влияния нервной системы на окраску кожи приводит следующего рода наблюдения. Человек год тому назад во время сильных болей приобрел темную пигментацию кожи (как у мулата) и почти одновременно с тем у него появилась параплегия нижних конечностей; при этом не замечалось ни явлений общей слабости, ни сильного душевного угнетения, что исключает возможность предположения о развитии бронзовой болезни. Другое наблюдение, сделанное тем же автором, относится к чернокожему, кожа которого на некоторых местах, как, например, в области бровей и на груди, сделалась белею. При этом не существовало никакой внутренней болезни, но замечалась необыкновенная слабость и упадок сил, как при бронзовой болезни. Кроме того, автор цитирует интересный случай Гэлла: женщина, страдавшая lepra anaesthetica, имела темную окраску кожи на местах анестезии. В той же статье приводится случай (опубликованный в Indian annales of med. science) анестезии пятой пары черепномозговых нервов, сопровождавшейся отложением пигмента в области ram. frontalis. Замечательно,, что при возвращении способности ощущения у больного снова восстановился нормальный цвет кожи. У помешанных, в особенности с симптомами мозгового возбуждения, Морселли 3 наблюдал особенную форму vitiligo, причем бесцветные пятна, резко пигментированные по окружности, захватывали симметричные участки кожной поверхности, преимущественно в области шейного симпатического нерва. Также у ипохондриков нередко встречают темную окраску кожи, которая, повидимому, не имеет никакой связи с расстройством отправления печени, а зависит, по всей вероятности, от нарушенной иннервации. Наконец, в последнее время были опубликованы весьма интересные случаи образования кожного пигмента (nigrities) у помешанных, в особенности при прогрессивном параличе 4. Су-
1 Britisch med. and foreign review, April 1861.
2 Barlow, Med. times and gaz., Nov. 1 1862: Schmidt's Jahrb., Bd. 126.
3 Мог sell i, Vitiligo des alienes, Rivista di freniatria, 1879, fasc. 12, p. 77.
4 См. случай Февра (Fdvre) в Annales med. psych., 1876, p. 26 et 197; 1877, p. 375.
188
ществуют также примеры аномалий кожного пигмента в течение различного рода чисто нервных расстройств. Так, местные обескрашивания кожи иногда наблюдались при существовании мозговых опухолей (случай Бурневиля) г. Дюмениль наблюдал изменения кожного пигмента при хроническом неврите. Куиба указывает на пигментацию, происходящую в направлении локтевого и срединного нервов при существовании гиперестезии в той же области 2. Деламар недавно описал случай травмы головы в лобно-теменную область. Вскоре после того у больного появилось темное пигментированное пятно соответственно foramen mentale с выпадением волос на правой брови отчасти на правом ухе и на подбородке с правой стороны, за исключением нижней его поверхности, обращенной к шее. Впоследствии присоединились явления прогрессивной атрофии, развились головные боли, и больной в конце концов помешался причем у него появилось другое пигментированное пятно с левой стороны, а также соответственно foramen mentale 3. Наконец, здесь следует упомянуть, что еще несколько лет тому назад Симон описал интересную форму naevus, названную им пае-vus nervosus, которая встречается большею частью на одной половине тела и располагается по ходу нервов 4. Подобные случаи наблюдались также Береншпрунгом, Томсоном, Рай-ером и др. Кроме того, во времени Бигеля стали известны случаи альбинизма и нигризма, ограничивающихся определенными нервными участками. Все эти случаи доказывают, что существует известное влияние со стороны нервной системы на образование и количественное отношение кожного пигмента8. Условия, при которых поражение нервной системы отражается на содержании кожного пигмента, нам пока еще малоизвестны, но нельзя не видеть здесь полной и несомненной аналогии с тем, что известно под названием питательных или трофических расстройств, в зависимости от поражений нервной системы. С этой стороны указанные факты имеют для нас живой интерес и побуждают к собранию наибольшего числа точных наблюдений, что заставляет меня опубликовать следующий случай.
Больная девица Н., 14 лет, для своего возраста малого роста, плохого сложения и мало упитанная. Родители больной в настоящее время живы и отличаются здоровьем; сифилиса и пьянства в семье не было. Мать имела роды 15 раз, но из всех детей только 7 родились живыми. Первые пять родов протекли нормальным образом; при следующей же беременности,
1 Bourneville et Poirier, Tumeur du lobe fronto-parie-tal gauche, Progres medical, 1879, N 24.
2 Arnozan, Des lesions trophiques, Paris, 1880, p. 153.
s Delamare, Virch. Iahresber., 1880, Art. Psychiatrie und Nervenkrankheiten, bearb. v. Westfahl.
4 Arch. f. Dermat. u. Syphilis, 1872.
5 В болезни, известной под названием бронзовой или аддисоновой, причина явления, по всей вероятности, заключается в подобном же влиянии со стороны нервной системы.
189
по счету шестой, мать была испугана и ребенок родился мертвым и недоношенным. Все следующие 5 детей родились также мертвыми недоносками. После того родилась наша больная доношенною; за год до этих родов мать перенесла тиф, а во время беременности страдала воспалением печени; в остальном была здорова, и роды, вообще говоря, протекли правильным образом. После того мать имела еще двух детей, которые в. настоящее время здоровы. Вообще все оставшиеся в живых дети теперь здоровы и не страдают никакими нервными явлениями. В детстве наша больная перенесла корь, коклюш и страдала рахитизмом с явлениями незначительной головной водянки. Затем до десятилетнего возраста оставалась почти совершенно здоровою и, по словам матери, развивалась в умственном и физическом отношении довольно правильно, но отставала от других своим ростом; только в последние годы рост ее значительно подвинулся вперед. Девочка всегда отличалась резвым и веселым характером. С давних пор у нее стали замечать неправильность стула, которая в последнее время начала усиливаться. Два года назад в первый раз, без видимой причины, начались частые головные боли; около того же времени мать заметила темножелтоватую окраску кожи, в виде пятна на спине, между лсттками. Пятно со временем стало разливаться, и темножелтоватая окраска распространилась в виде пятен на ббльшую часть спины, живота, а затем и на другие части тела. Среди этой пигментированной кожи местами выделялись пятна, резко отличавшиеся своей белизною. В результате иа некоторых частях тела, в особенности на животе, получилась картина пестрой «тигровой кожи», как ее называли домашние. Кроме того, вместе с развитием головных болей у девочки обнаружилась седина среди ее черных густых волос, сопровождавшаяся выпадением их. Желтоватая окраска кожи, между тем, постепенно делалась более и более интенсивною, в особенности на животе, груди, спине, шее, и в последнее время захватила ббльшую часть тела. Зубы также пожелтели и затем многие из них искрошились. Белки глаз и ногти всегда оставались с нормальной окраской. Вместе с развитием болезни девочка начала худеть, сделалась вялою, а в последнее время раздражительною. От умственных занятий она была освобождена по совету врачей и об успешности их мать не может сказать ничего определенного. Месяца три назад присоединились нервные расстройства: больная стала испытывать парестезии, чаще всего в виде ползания мурашек в кистях обеих рук, также в щеках и в кончике носа, вскоре затем время от времени начали появляться настоящие судороги и контрактуры в пальцах обеих рук, вследствие чего некоторое время больная не могла их ни разогнуть, ни согнуть и совершенно лишалась свободного употребления конечностей. Описанные явления судорог и контрактур, всегда начинаясь ощущением мурашек, наступали почти ежедневно, без исключения, и продолжались по нескольку часов кряду, а в последнее Время иногда даже по целым дням. Ночью, когда больная просыпалась, она тоже часто жаловалась матери на судороги в пальцах. Кроме того, в последнее время стали замечать судорожные подергивания в губах и дрожание век. Головные боли в настоящее время повторяются довольно часто; попрежнему постоянные гастрические расстройства, слабит чрезвычайно жидко. Тошноты и рвоты никогда не замечалось. Общая слабость заметно усиливается; больная легко устает при ходьбе и при всяком физическом усилии; умственные занятия служат ей в тягость. Сон большей частью не совсем спокойный и сопровождается грезами.
При исследовании на амбулаторном приеме в клинике в первых числах апреля месяца найдено следующее: больная представляется недостаточно развитою соразмерно своему возрасту. Рост ее едва превышает 2 аршина; телосложение вообще слабое, слизистые оболочки бледны, вся мышечная система в плохом развитии; грудные железы малы и не вполне сформировались. Регул до сих пор еще не появлялось. На лице обращает на себя внимание общий бледновато-желтый оттенок кожи, в особенности заветный под глазами, около носа и в области верхней и нижней губ. Кожа лба представляется желтого цвета, местами с белыми пятнами,
190
лишенными пигментов; шея пигментирована гораздо сильнее: вся задняя часть ее, отчасти и передняя, темножелтоватой окраски, также с белыми пятнами. При обнажении тела оказывается, что подобная окраска имеет весьма обширное распространение. Вся кожа спины, также груди, обоих боков и в особенности поясничной области и живота имеет сплошную тем-ножелтую окраску, за исключением нескольких, различной величины, кругловатого очертания пятен, лишенных пигмента; плечи также пигментированы, но слабее; на предплечьях же и кистях рук пигментации или вовсе не замечается, или только весьма слабая. Эта окраска не изменяется при давлении пальцем и не исчезает при поверхностном царапании: при этом никакого шелушения и вообще видимых изменений в ткани самой кожи не замечается. Вниз темножелтоватая окраска распространяется до колен; кожа обеих голеней представляется бледнорозового цвета; на ступнях же снова замечается явственная желтоватая окраска. При исследовании полости рта оказывается, что спинка языка и в особенности, края его пигментированы в розоватожелтый цвет; кроме того, в языке замечаются слабые фибриллярные сокращения мышц. Из зубов верхние резцы совершенно испорчены, так же как и многие из коренных зубов; нижние резцы совершенно желтые, и мать уверяет, что они выпадут, так как верхние сначала также пожелтели, а затем начали крошиться. Белые пятна, о которых сказано выше, в особенности распространены по коже передней и задней стенки груди и на животе; здесь они достигают от едва заметной величины до 2 — 3 пальцев в поперечнике. По большей части эти пятна имеют симметричное распределение. Так, замечается несколько маленьких пятен на обеих сторонах шеи среди темножелтоватой окраски и по одному большому пятну в обеих подмышечных областях; кроме того,, оба соска представляются совершенно обескрашенными; затем несколько различной величины пятен встречается на обеих сторонах задней части грудной клетки, в особенности в области лопаток; далее — 3 пятна, величиной с копейку, симметрично расположены по обеим сторонам Ппеа alba, на боковых стенках живота; по одному большому пятну (в ладонь величиною) замечается в области больших вертелов и, наконец, несколько маленьких пятен в обеих подколенных ямках. Почти все остальные пятна расположены по средней линии тела преимущественно в верхней части позвоночника, также по sternum, вдоль linea alba и одно пятно в области pubis. На некоторых из этих пятен почти вовсе не замечается признаков существования волос; на других только весьма слабое проявление светлого пушка. Чувствительность, как на местах пигментированных, так и лишенных пигмента, не обнаруживает резких изменений. На волосистой части головы обращает на себя внимание довольно значительная седина среди темных волос. Кроме того, на макушке головы заметна плешина, величиною с двугривенный, появившаяся недели две тому назад; другая, меньшей величины, плешинка расположена в середине теменной области. Кожа на местах плешин совершенно чистая, гладкая; матово-белая, не шелушится и с нормальной чувствительностью. В других частях головы волосы довольно редкие. Подмышками также замечается несколько поседевших волос. Мне остается прибавить, что кожа у нашей больной повсюду сухая и вялая; подкожный слой жира чрезвычайно тощий. Общая худоба в особенности резко обнаруживается на туловище: ключицы, ребра и лопатки обрисовываются рельефно; на реберных хрящах прощупываются четкооб-разные утолщения. Сухожильные и мышечные рефлексы повсюду нормальны. При исследовании внутренних органов не замечено никаких патологических изменений ни в грудных, ни в брюшных органах, в области живота никакой болезненности. Пульс большей частью слабый, около ста ударов в 1 минуту. В моче отсутствие белка и сахара. Назначено: мышьяк с железом в виде solut. arsen. Fowled cum tinct. tonicae Bestuch., бромистый калий и питательная диэта.
23 апреля — судороги и контрактуры в пальцах не повторялись уже с неделю; также последние дни не было ощущений мурашек в кистях рук,. но еще бывают на несу и щеках. В последние дни нередко случались голов-
191,
ные боли. Сон попрежнему с грезами, но крепкий. Во время амбулаторного приема иногда замечаются дрожание век и судорожные подергивания губ.
8 остальном все попрежнему.
25 апреля. В прошедший раз больная на приеме за короткое время утомилась настолько, что еще дорогой начала испытывать большую наклонность ко сну. Затем в тот же день к вечеру появилось колотье в правом боку, которое, впрочем, скоро прекратилось само собою. Ни судорог, ни ощущения мурашек до сих пор не появлялось. Окраска кожи в прежней степени, но белые, не пигментированные пятна в некоторых местах заметно расширились.
5 мая. За последнее время у больной развилась otitis externa subacute в обоих ушах, замечается незначительная течь из левого уха. Больная держится бодрее прежнего; раздражительности не замечается.
9 мая — девочка жалуется на головную боль, вялая и скучная. По словам матери, вчера она утомилась в игре со своими товарками. Снова замечается раздражительность и по временам ощущение мурашек в руках. Течь из левого уха продолжается. Слух с этой стороны немного ослаблен.
12 мая. Больная снова с головной болью, по временам появляется ощущение мурашек в пальцах рук. Сегодня во время снятия гипсового слепка со стана девочки у ней появились кратковременные судороги в пальцах, захватывающие преимущественно экстензорные мышцы предплечья.
В течение второй половины мая месяца больная почти все время жаловалась на головную боль. Кроме того, время от времени у девочки появлялись приступы судорог в обеих руках и ощущение мурашек как в руках, так и в лице. Под влиянием нескольких приемов хинина все эти явления начали снова уменьшаться. Пигментация кожи у больной в прежней степени; на местах же головы, лишенных волос, появился довольно густой белый пушок. Неправильность стула попрежнему. Все лето больная провела на даче, исполняя соответствующие указания терапии (приемы железа, мышьяка, хинина и питательная диэта). Я мог видеть ее снова только
9 сентября. Больная в это время представлялась пополневшей; вес ее тела увеличился на несколько фунтов. Сама она чувствовала себя гораздо бодрее сравнительно с прежним временем; угнетенного состояния давно уже не замечалось. Головные боли также прошли; ни ощущения мурашек, ни судорог больная не испытывала уже в течение около двух месяцев; желудок исправился почти совершенно, так что больная считает себя значительно поправившеюся. Пигментация кожи почти в прежнем положении, но только интенсивность окраски сделалась немного слабее. Точно так же области, лишенные пигмента, остались почти в том же положении или только немного расширились. Пигментация слизистой оболочки языка выражена еще в значительной степени и зубы попрежнему остаются интенсивно желтыми. На голове седина значительно увеличилась, в особенности в лобно-теменной и височной областях, а бывшие две плешинки совершенно заросли нежными белыми волосами. В последнее время больная имела в первый раз регулы, протекшие совершенно нормальным образом.
Как мы видели, у больной Н. существовал совершенно особенный симптомокомплекс нервных явлений, выражающихся ненормальной раздражительностью, общей слабостью, головными болями, судорогами и парестезиями в конечностях вместе с нутритивными изменениями общих покровов —усиленной пигментацией кожи и слизистых оболочек рядом с местной атрофией пигмента, преждевременным поседением и облысением головы на известных местах. Случай этот нас интересует в данную минуту главным образом в отношении рас-
.192
стройств питания кожи и прежде всего в отношении аномалии кожной пигментации. Здесь мы имеем как распространенную гипертрофию кожного пигмента, более или менее разлитую, так и местную атрофию его (vitiligo). Известно, что в обыкновенных случаях гипертрофия кожного пигмента может происходить: 1) как последствие различного рода сыпей, в особенности сопровождающихся зудом (вследствие чесания); 2) пигментация кожи усиливается под влиянием некоторых раздражителей, действующих непосредственно на кожу (солнечные лучи), и, наконец, 3) развитие пигмента находится в некотором отношении с известными физиологическими состояниями организма, как, например, с развитием половой деятельности у женщин (так называемые печеночные пятна или chloosmata uterina). О первых двух из этих источников образования кожного пигмента по отношению к нашей больной не может быть и речи. Что касается третьего, то, помимо того, что chloosmata uterina, локализируясь преимущественно на лице, почти никогда не достигают столь обширного распространения, как у нашей больной, необходимо еще иметь в виду, что появление этих пятен обыкновенно находится в известном отношении с развитием половой сферы, тогда как наша больная при начале болезни не имела даже менструаций.
Итак, в данном случае мы не находим другого источника для развития ненормальной пигментации кожи и слизистой оболочки языка, помимо нервной системы, влияние которой в этом отношении, как мы видели на многих цитированных случаях, может быть доказано с очевидностью. Следовательно, ненормальную пигментацию покровов у нашей больной мы рассматриваем как трофоневроз. Справедливость этого взгляда, с одной стороны, подтверждается также существованием различного рода нервных симптомов у нашей больной, с другой стороны —одновременным появлением других, несомненно, трофических, расстройств, к рассмотрению которых мы сейчас перейдем *.
1 Здесь необходимо сказать, что понятие о трофоневрозе в отношении к аномалиям кожного пигмента в настоящее время не может считаться лишенным патологоанатомической основы. Мы подразумеваем в данном случае недавние исследования Лелуара (Arch, de physiologie normal, et patholog., 1881, N 3, p. 391), который в двух случаях vitiligo (наблюдавшимися также и в нашем случае) находил кожные нервы измененными: волокна их представляли с большой ясностью явления атрофического неврита; в некоторых из них осевой цилиндр совершенно исчез, миэлин представлялся разломанным и в каплях, кроме того, замечалось размножение ядер, и шванновское влагалище содержало вещество, окрашивающееся пикрокармином в желтый цвет, которое обыкновенно встречается в нервах, подверженных регенерации. На основании этих положительных данных, автор присоединяется к взгляду Алибера, Райера, Гебра, Робина, Неймана, Дюжарден-Бомеца, Февеоля, Морселли, Бурневиля, Пуарье, Бьюкли, Гаскойна и Висса, считающих vitiligo заявление трофического свойства.
13 В. М. Бехтерев
193
III. Преждевременное поседение и выпадение волос
В приведенном наблюдении, как мы видели, кроме аномалий кожного пигмента, замечалось еще преждевременное поседение волос, сопровождавшееся местным выпадением их. Явление это, наступившее в нашем случае почти одновременно с развитием головных болей и других нервных симптовов, как мы уже сказали, принадлежит к числу трофических изменений. В литературе вообще существует немало примеров относительно нутритивных изменений волос при различных патологических процессах в нервной системе. Так, при травматических невралгиях, преимущественно лицевых нервов, нередко замечают большую их ломкость, грубость и весьма часто особенную болезненность. В других, менее частых, случаях наблюдали изменение цвета волос. Известен пример Энети, который на себе самом наблюдал, что каждый раз во время приступа надглазничной невралгии волосы соответственной брови с той же стороны седели. Это изменение цвета продолжалось всегда в течение нескольких дней и затем волосы снова принимали прежний цвет (без малейшего выпадения) г. Гюблер и Бордье наблюдали замечательный случай местного обесцвечения волос на голове во время приступа мигрени, которое впоследствии еще более усилилось. По истечении нескольких месяцев, на большей части головы больного можно было наблюдать вперемежку поясы белых и черных волос 2. Но еще более поразительными представляются те случаи, когда изменение цвет?! волос наступает быстро, под влиянием сильного нервного потрясения, например, страха 3. Весьма интересным представляется также случай Целера —с периодическим душевным расстройством, где во время отдельных приступов болезни цвет волос менялся с черного на серебристый 4. Ландау описывает другой случай, где у больного, страдавшего белою горячкою, уже на второй день после приступа буйства темные волосы поседели. Выпадение волос точно так же наблюдали в таких случаях, где связь с поражением нервной системы представлялась
1 См. Эйленбург, Руководство к нервным болезням, в отделе о мигрени.
2 G О b 1 е г et Bordier, Art. Migraine, Diet, encycl., t. VII, 2 serie, p. 630.
3 См. интересные случаи, собранные Пуше в Revue des deux mondes, Janvier 1872.
Капози (Патология и терапия накожных болезней, Харьков, 1881) в недавнее время высказал сомнение в возможности существования подобного рода случаев потому только, что он считает их невозможным для себя удовлетворительно объяснить. Между тем факты такого рода были засвидетельствованы такими авторитетными и осторожными наблюдателями, как Шарко (De la conitie rapide, Gaz. nebdom., 1861), Пинкус (Virch. Arch., Bd. 43, 44), и др.
4 См. Ill ю л е, Руководство Цимсена, т. 16, стр. 156.
194
очевидною. Так, довольно обыкновении случаи выпадения волос вследствие травматического неврита или при невралгиях, в особенности на конечностях. В таких случаях выпадение волос почти всегда бывает временным и исчезает вместе с прекращением нервного страдания г. Описаны также, случаи выпадения волос при травматических повреждениях спинного мозга на всей парализованной поверхности тела 2. О случае Деламара, где выпадение волос осталось последствием травмы головы, было уже сказано выше. Здесь следует упомянуть еще, что Раватон вслед за сотрясением головы наблюдал развитие амавроза на одном глазу и выпадение волос с головы, бровей и ресниц на больной стороне. Ромберг приводит случай одностороннего выпадения волос при paralisis facialis. Далее всем известно, что под влиянием угнетающих аффектов, тяжелых душевных испытаний, забот и также вследствие усиленной умственной деятельности весьма нередко наступает преждевременное выпадение волос. В этом отношении Фреде опубликовал замечательное наблюдение одной девочки, у которой вслед за внезапным испугом в течение несколько дней выпали волосы не только на голове, но также и на теле. Данные, подобные предыдущим, были подтверждены также и экспериментальной физиологией. Известно, что перерезка седалищного нерва у животных почти всегда вызывает последовательное выпадение волос, которые, впрочем, снова вырастают как в случаях, где нерв возрождается, так и в случаях противоположных. Наконец, выпадение волос в болезни, известной под названием alopecia areata s. area celsi (la pelade), наблюдавшейся также у нашей девочки, из-давна считалось большинством дерматологов за явление трофического свойства, вследствие расстройств нервной системы, В недавнее время, впрочем, явились некоторые опровержения (Бюхнер, Гарди) упомянутого взгляда в пользу паразитической теории, —опровержения, до сих пор, однако, не имеющие твердой фактической почвы. Во всяком случае, факты, когда выпадение волос без всякой внешней причины и воспалительных явлений в коже наступает рядом с развитием или в течение нервных расстройств, сохраняют свою положительную силу и значение в смысле трофических изменений, тем более, что влияние нервной системы на выпадение волос, как мы видели, подтверждается также экспериментальной физиологией. Для иллюстрации я желал бы привести здесь еще один поразительный по своим последствиям случай выпадения волос (alopecia) с ясной этиологией со стороны нервной системы 3.
1 Poinsot (cit. par Arnozan), Des lesions trophiques, 1880, p. 155.
a Jelly, Britisch. med. journ., 1873.
8 Случай этот в течение прошедшего года можно было наблюдать в клинике профессора Полотебнова,_ куда больной был впоследствии помещен. Подробное исследование его было призведено там же.
13* 195
Мальчик крепкого сложения, довольно полный на вид, 11 лет, был приведен в конце мая месяца нынешнего года в Георгиевскую общину для совета с доктором Томашевским относительно общего выпадения волос. Из рассказа отца можно узнать следующее. Мальчик был блондин. Волосы начали выпадать с великого поста прошедшего 1880 года. Раньше мальчик почти никогда ничем не страдал; только четыре года тому назад он перенес какую-то лихорадочную болезнь, сопровождавшуюся сыпью по телу, но волосы после нее не падали и оставались совершенно крепкими, как и прежде. Ушибов в голову никогда не было. Недели за четыре до выпадения волос он изредка начал жаловаться отцу на головные боли и частые головокружения. Сам больной в настоящее время, впрочем, отрицает головные боли, но говорит, что в течение всего великого поста страдал чрезвычайно сильными головокружениями. По его словам, вдруг вокруг него все закружится; его самого начинает шатать из стороны в сторону, иногда он падает и на некоторое время теряет сознание. Такого рода припадки с ним происходили почти каждый день в течение нескольких недель. Вскоре начали выпадать волосы целыми прядями, и в короткое время выпали совершенно. Несколько времени спустя на гладкой поверхности головы появился темный пушок, но затем снова выпал совершенно. Позднее начали выпадать волосы на бровях, подмышками и на других частях тела. Почти одновременно с этим появились парестезии в области глаз, в виде ощугцения холода, — как он выражается «глаза как будто мерзли», на что больной нередко жаловался своему отцу.
Во время исследования кожа головы представлялась совершенно гладкою и чистою, без всякого следа волос; ни малейшего признака воспалительных явлений на ней не замечается. Точно так же на бровях, подмышками, в области genitalia и вообще на всей поверхности тела нельзя различить ни одного волоска, за исключением ресниц, которые остались почти в целости. Других каких-либо аномалий не замечено. Подробное исследование чувствительности не указало никаких патологических уклонений.
Случай этот, помимо своей редкости, в особенности интересен в отношении своей этиологии. Как мы видели, кроме чисто нервных расстройств, выражавшихся головными болями и весьма сильными и частыми головокружениями в течение многих недель, нельзя было отыскать у нашего больного никакой причины для наступившего выпадения волос. Будущее, может быть, разъяснит нам как значение вышеуказанных этиологических моментов в подобных случаях, так, может быть, и точнее укажет патологическую основу рассматриваемого страдания.
ОДЕРЕВЕНЕЛОСТЬ ПОЗВОНОЧНИКА С ИСКРИВЛЕНИЕМ ЕГО КАК ОСОБАЯ ФОРМА ЗАБОЛЕВАНИЯ
В течение последних лет я имел возможность наблюдать заболевание, выражающееся в развитой форме искривлением кзади и неподвижностью или одеревенелостью позвоночного столба и целым рядом расстройств со стороны нервной системы. Так как при обзоре подходящей литературы я не нашел более или менее подробных и вообще точных указаний по поводу этого своеобразного страдания, то, интереса ради, я и считаю не лишним сообщить свои наблюдения по сделанным мною записям о больных этого рода, из которых трое были пользуемы мною на домашнем приеме.
I. Больная Ш., 56 лет, явилась ко мне на прием с жалобами на мучительное чувство тяжести в задней части шеи, отдающееся в спинной хребет и в заднюю часть головы; кроме того, по временам больная испытывает ощущение стягивания в горле и ее как бы душит. Более же всего она жалуется на то, что за последнее время ей стало трудно поддерживать голову, которую постоянно клонит вперед. С первого же раза у больной обращает на себя внимание резкое искривление позвоночника кзади в верхней его части, в особенности в нижней шейной и лопаточной областях, причем голова представляется как бы выдвинутой вперед и слегка опущенной. Мать больной тоже страдала искривлением позвоночника кзади, хотя и более, слабым; затем подобным же искривлением позвоночника страдала сестра больной и, кроме того, с детства страдает им и ее дочь.
Сама больная была несколько сутуловата еще с молодости, но в пос-следние годы искривление позвоночника, как она замечает, стало резко усиливаться. Вместе с этим усилением искривления вначале были боли в правой руке; кроме того, в течение последних 5 лет больная нередко испытывала чувство тяжести в голове, которое усилилось в особенности за последнюю зиму. Теперь тяжесть обнаруживается преимущественно в затылочной области головы и в задней части шеи. Сверх того, в последнее время больную стало постоянно беспокоить своеобразное и тягостное чувство стягивания в боковых частях шеи, преимущественно под нижней челюстью, в особенности слева. Из других явлений следует отметить, что прежде у больной бывали сильные мигрени. Лет 20 назад она мгновенно ослепла на правый глаз, но затем зрение ее стало возобновляться, хотя еще и в настоящее время правый глаз видит хуже левого. Явлений истерии не было никогда.
Status praesens. Выпуклая горбатость (искривление кзади) позвоночника в нижней шейной и верхней грудной областях, благодаря чему голова сильно выдвинута вперед. Лордоз в нижней части позвоночника если и есть, то весьма незначительный. При более внимательном осмотре тулс-
197
вища замечается лишь некоторое опущение лопаток и как бы расхождение их в стороны, но ясной атрофии мышц, поддерживающих лопатки, под жировым покровом незаметно; только на шее справа заметно некоторое исхудание мыши по сравнению с левой стороной. Болевая чувствительность в лопаточных областях спины незначительно понижена; никаких других расстройств чувствительности нет, кроме упомянутого выше чувства стягивания под нижней челюстью и ощущения тяжести в задней части шеи и затылочной области головы. При выпрямлении, т. е. поднятий головы кверху, чувство тяжести в ней всегда усиливается. Точно так же, по заявлению больной, чувство тяжести заметно усиливается и при ходьбе. Самородных болей в спине больная не испытывает; при надавливании на позвоночник и при постукивании по нем боли тоже не ощущает. Органы чувств в порядке, за исключением ослабления зрения на правый глаз, которым видит неясно, причем верхние части предметов кажутся как бы затянутыми легкой пеленой. В области движения главная жалоба больной сводится на то, что голову тянет вперед, так что трудно поддерживать ее в естественном положении. Выпрямлять голову ей тоже трудно; И, как я уже сказал, при этом всегда является чувство тяжести в задней части шей и головы. Движения шеи в стороны довольно свободны. Тип дыхания исключительно брюшной: плоская грудь больной почти вовсе не участвует в дыхательных движениях. Позвоночник* в грудной области поразительно мало подвижен. Повороты в сторону, выпрямление и сгибание его чрезвычайно ограничены В лежачем положении горбатость не исправляется. Других двигательных расстройств незаметно. Сухожильные рефлексы без существенных изменений. В течение последнего года у больной стали обнаруживаться запоры. В остальном никаких расстройств. Лечилась электричеством, но улучшения не было; скорее даже от электричества ей стало хуже.
Во время наблюдения, продолжавшегося около месяца, больная принимала йодистый калий, а против болезненных ощущений —временам хинин с антифебрином. Кроме того, делались прижигания по позвоночнику в шейной области. Прочного улучшения не последовало, но прижигания, несомненно, облегчали, хотя и временно, тягостные ощущения стягивания в шейной области.
II. Больная М., 52 лет, обратилась ко мне за советом в начале августа 1890 г. по поводу приступов сердцебиения, тоски и бессонницы, которыми она страдает уже несколько месяцев. Резкое искривление позвоночника кзади, преимущественно в верхней части грудинной области. Из расспросов по этому поводу выяснилось, что она уже более 10 лет стала замечать, что верхнюю часть ее туловища постепенно клонит вперед. Незадолго перед этим (лет 12 назад) больная упала в бане и сильно ушибла спину, которая после того была черного (кровоподтек). Вслед за тем больная стала испытывать боль в нижних шейных позвонках, если долго оставалась в сидячем положении. В то же время стала обнаруживаться впервые небольшая сутуловатость. Эти болезненные явления сама больная относила к тому, что, живя без прислуги, она должна была много хлопотать по хозяйству, причем часто приходилось работать в согнутом положении. Два года назад, отворяя дверь, она случайно упала на спину и ушибла себе руку, после чего почувствовала боль в спине с месяц и должна была лежать в постели. От боли она не могла даже повертываться в кровати без посторонней помощи; кроме того, полученные при падении повреждения руки осложнились рожею. Во время лежания в постели у больной появились месячные, которых ранее не было уже 8 лет. Через месяц больная поправилась настолько, что при лежании уже не чувствовала боли в спине, но при ходьбе и сидении боль еще была. Эта боль в меньшей степени продолжается и по сие время, вследствие чего больная и не может подолгу сидеть. Со времени падения сутуловатость стала быстро усиливаться до резкегэ искривления позвоночника кзади, причем это искривление, по заявлению больной, и в настоящее время постепенно усиливается все более и более. Из анамнеза больной выясняется далее, что ее мать тоже стра-
198
дала искривлением позвоночника кзади, но в меньшей степени, тетка же больной страдала подобным искривлением в крайне резкой степени. Больная заявляет, что в настоящее время она чувствует по временам жар в теле, не исключая и головы, сопровождающийся потением. Это явление обыкновенно наступает внезапно, в различное время дня и, продолжаясь не более 10 минут, сопровождается чувством общей слабости в такой степени, что больная в это время не может сидеть. Только что указанное явление в особенности стало обнаруживаться после второго падения; теперь оно бывает иногда по нескольку раз в день. После второго падения у больной бывает также сильная бессонница и тоска. Что же касается до сердцебиения, то больная! страдает им уже с давних пор. Следует еще упомянуть, что в руках нередко появляется чувство онемения, а по временам бывает и дрожание; в последнее время в руках стала обнаруживаться даже некоторая слабость. Кроме того, больная чувствует, как будто голову €е гнет вперед, так что нужно некоторое усилие, чтобы поддержать ее в прямом положении.
Первое, что бросается в глаза, — это упомянутое уже резкое искривление позвоночника, преимущественно в верхней грудной области. При осмотре туловища сбоку можно сказать, что верхняя часть позвоночника образует выпуклую кзади дугу с радиусом приблизительно в 2/4. Вместе •с тем голова больной заметно выдвинута вперед, шея кажется укороченной, подбородок же слегка опущен. В поясничной области ясного уравновешивающего лордоза нет. Таз стоит более прямо и как бы несколько выдвинут вперед; расстояние же между мечом и лобком укорочено, причем живот как бы выпячен вперед, тогда как грудь, наоборот, предствляется плоскою, почти впалою. Выпрямление позвоночника невозможно ни при каких условиях, даже и при лежании. Небольшое наклонение туловища вперед и назад возможно лишь благодаря некоторой сгибаемости позвоночника в нижней грудной и поясничной частях. Повороты позвоночника возможны также лишь в слабой степени, благодаря подвижности его нижней части. Верхняя же половина грудной и нижняя часть шейной областей позвночника почти совершенно неподвижны. При попытках к поворачиванию туловища чувствуется некоторая боль в шейной области. Поднятие головы кверху и повороты ее в стороны заметно ограничены; последние совершаются почти исключительно за счет сочленения атланта со вторым позвонком, вследствие чего движение головы делает впечатление флюгера, движущегося на неподвижной оси. На спине заметна некоторая атрофия мышц в надлопаточных областях. Подостные мышцы тоже слегка атрофированы, в особенности правая. Шея и руки кажутся несколько исхудавшими. Реберные промежутки почти всюду ясно очерчены. Лопатки и наружные части ключиц сильно опущены, благодаря чему плечи стоят несколько ниже обыкновенного. Чувствительность всех родов, не исключая и болевой, ясно понижена на спине, в особенности соответственно уровню первых 4 грудных нервов. В меньшей степени понижение чувствительности обнаруживается и в верхней половине груди, а также в надключичных областях шеи; в нижней части груди ясного понижения чувствительности нет. Из субъективных расстройств чувствительности обращает на себя внимание по временам бывающее чувство ползания мурашек по спине и онемение в руках. Кроме того, у больной уже с давних пор по временам бывают боли в ногах, в особенности в подколенных областях и в вертлугах. Органы чувств в порядке. В позвоночнике самородных болей не чувствует вовсе; лишь при сильном постукивании в области 4 верхних грудных позвонков обнаруживается некоторая болезненность, остальные же позвонки и при постукивании безболезненны. В области движений обнаруживается уже упомянутое ослабление силы рук. Хотя все движения рук и возможны, но движения их, в особенности вверх, за голову и кзади, за спину, крайне слабы и вызывают обыкновенно общую усталость. Фарадическая и гальваническая возбудимость мышц не изменена; лишь в мышцах лопаточных областей заметно некоторое ослабление электрической возбудимости. В походке заметных расстройств
199
нет. На левой ноге два пальца согнуты и обнаруживают неподвижность суставов. Дыхание исключительно брюшное: грудная клетка почти вовсе не участвует в дыхании. Сухожильные рефлексы без существенных изменений, но при ударе молоточком по внутреннему краю нижнего угла лопатки не получается отраженного движения руки, бывающего у людей здоровых. Мочевой пузырь работает совершенно правильно. С давних пор больная страдает запорами. В матке — полип. В остальном ничего патологического.
За время наблюдения, продолжавшегося около 3 недель, больная принимала против тоски и сердцебиения бромистый натрий с кодеином и настойкой ландышей, а против бессонницы и запоров — бромистый хинин с ревеием; кроме того, были назначены общие спиртные растирания, тепловатые ванны в 28°Р с солью и прижигания по позвоночнику в верхней грудной области. Под влиянием этого лечения тоска стала значительно меньше, серцебиение совершенно прекратилось, бессонница прошла, запоры уменьшились. Неповоротливость же спины и все другие явления остались без изменения.
III. Больной Г., 39 лет, много раз падал и ушибал спину, 15 лет тому назад получил ушиб вьюком в левое плечо, так что упал и не мог сам подняться. Кроме того, нужно упомянуть, что, служа с 1870 по 1888 г. составителем поездов, больной часто должен был работать с наклоненным туловищем. Мало-помалу у него стало развиваться искривление позвоночника кзади, а несколько позднее, лет 10 назад, впервые появились и боли в туловище, руках и ногах. Боли эти частью были стреляющие и блуждающие, а частью сосредоточивались в локтевых и бедренных сочленениях, вследствие чего и признавались врачами за ревматические. Вследствие боли в суставах больной года \г/2 ходил с трудом. Спустя год появились и поясничные боли, затем года четыре спустя к поясничным болям присоединились боли в боках, столь сильные, что больной почти не мог ездить на извозчиках вследствие тряски. Кроме того, временами бывали парестезии в подошвах в виде щекотания.
Так как все боли, как упомянуто, признавались за ревматические, то в 1889 г. больной врачами был послан из Казани в Астрахань на Тина-кские грязи, но, как и следовало ожидать, никакого улучшения на грязях не последовало; скорее даже боли обострились и общее состояние ухудшилось. Между прочим, после Тинакских грязей появились приступы сердцебиения и боли в грудной доске и в левой половине груди, продолжающиеся и по сие время. Суставные боли были еще прошлой зимой, но облегчились от горчичников и в настоящее время бывают лишь изредка и на минуту, ие более. Чаще всего больного беспокоят теперь боли в пояснице и в боковых частях живота, бывающие приступами на довольно продолжительное время (до нескольких дней). Слета прошлого года,т. е. с год i:a-зад, больной стал замечать общую слабость, выражающуюся усталостью при ходьбе.
Мать больного, которой в настоящее гремя 70 лет, за последние 15 лет сгорбилась, подобно сыну, причем и у нее по временам бывают бели в ногах и чувство щекотания в подошвах; отец больного, повидимому, в чахотке; о других родных больной не межет дйть точных сведений. Status pracsens 12 мая 1891 г.; больной среднего роста, довольно порядочного сложения, с умеренным развитием подкожного жира. Рсзгая горбатость спины с выпуклостью кзади, преимущественно в лопаточной области; вследствие этого голова, благодаря уравновешивающему отклонению шеи вперед, кажется выдвинутой кпереди и слегка опущенной. В поясничной области ясного лордоза нет; скорее можно бы говорить об уплощении поясничной части хребта. Таз как бы несколько выдвинут вперед, и потому живот кажется выпяченным. Передняя часть груди резко уплощена и даже кажется несколько вдавленной. Лопатки заметно выстоят кзади Еследхт-вие исхудания мышц. Дыхание исключительно брюшное, г следствие чего живот в верхней его части постоянно втягивается и выпячивается. Грудная же клетка придыхгнии остается неподвижной; даже и при уст ленном вды-
200
хании она не принимает заметного участия в дыхательных движениях.. Вместе с тем обращают на себя внимание поразительная неподвижность-позвоночника, который не сгибается ни взад, ни вперед, ни вправо, ни влево. При попытках согнуться больной лишь слегка сгибает туловище. Движения головы кверху ясно ограничены. В лежачем положении разгибания позвоночника не происходит. По заявлению больного, сгибать спину ему что-то не дает, несмотря на то, что при попытках к сгибанию особенной боли в позвоночнике он не чувствует. При постукивании позвоночника особенной болезненности не обнаруживается; лишь в нижней грудной области чувствуется при этом некоторая боль в глубине. Больной заявляет, что его туловище как бы клонит вперед и что ему трудно выпрямиться и приподнять голову. Вместе с тем он жалуется на общую слабость и слабость ног в частности. Хотяпоходка его и не представляется резко расстроенной, но, вследствие неподвижности позвоночника при ходьбе и стоянии, колени подгибаются, а потому и самая походка кажется совершенно своеобразной. Благодаря той же неподвижности позвоночника и частью вследствие слабости ног больному трудно подниматься с постели и даже со стула; при этом больной обыкновенно помогает себе руками. В особенности трудно больному вставать после долгого сидения. При этом вставании почти всегда появляется боль то в пояснице, то в боках. Мышцы рук и ног худы и дряблы на ощупь.Болевая и осязательная анестезии на спине, начиная с области II грудного и до области нижних поясничных нервов, но на уровне XI — XII грудного нерва чувствительность значительно лучше, нежели в выше- и нижележащих областях, и может даже считаться повышенной. При этом на левой стороне спины чувствительность вообще притуплена более, нежели на правой. На ягодицах и в нижних конечностях чувствительность сохранена; в плечевых же областях замечается некоторое притупление болевой чувствительности. На груди ниже области II грудного нерва хотя и обнаруживается анестезия, но в значительно меньшей степени, нежели на спине. На животе нижняя граница анестезии определяется линией, идущей над остями подвздошных костей, ниже пупка приблизительно пальца иа три.
Из субъективных расстройств обращают на себя внимание: почти постоянное чувство как бы электризации в шейной области, направляющееся от головы вниз, затем ощущение тяжести в груди («как бы доска положена на грудь») и чувство опоясывания, появившиеся за последнее время. Креме того, во время сидения и лежаиия больной испытывает почти постоянное неприятное щекотание в подколенных областях, при усилении которого он невольно делает иногда дергающее движение негой (судорожное сокращение).Подобное же щекотание ощущается в локтях, причем больной нередко, чтобы облегчить себя, тоже производит дергающие движения рукой. Далее, у него «можжат» голени, в особенности после ходьбы. С другой стороны, при сидении или неловком шаге чувствуется некоторая боль в области бедренного нерпа под пупартоьой связкой, с той или другой стороны, но более с левой. При давлении в этой области, действительно, обнаруживается болезненность. Позвоночник при постукивании безболезнен, кроме области, соответствующей уровню XI и XII позвонков, где сильное постукивание сопровождается ясною болью. На этом же уровне чувствуется глубокая боль и при давлении по обеим сторонам позвоночника, причем всегда является и потребность вдохнуть или задержать воздух в груди, как бы для облегчения болезненного ощущения. Точно так же чувство боли в вышеуказанных областях и та же своеобразная потребность задержать воздух в груди появляются и при езде на извозчике. Органы чувств без изменений; 'зрачки несколько неравномерны, но на свет реагируют правильно. Деятельность мочевого пузыря не нарушена. С некоторых пор заметна половая слабость. Коленные рефлексы на обеих сторонах повышены.
Во время наблюдения, продолжавшегося около недели, больной принимал йодистый калий и хинин с фенацетином. Кроме того, производились прижигания по позвоночнику прибором. Паклена. Существен-
201
''лого улучшения за такой короткий срок, конечно, последовать не могло, но прижигания, все-таки, несомненно, на некоторое время облегчали признаки раздражения чувствительных нервов.
В дополнение к приведенным наблюдениям считаю необходимым заметить, что еще прежде я наблюдал 2 больных с подобными же явлениями, но, к сожалению, ке сохранил о них подробных записей. Тем не менее,, в связи с предыдущими наблюдениями, и эти случаи не лишены интереса.
В одном из них у душевнобольного более 10 лет существовала выпуклая горбатость позвоночного столба в плечевой и шейной областях, достигавшая такой степени, что подбородок почти прикасался к грудной доске. При этом совершенно безболезненный позвоночный столб отличался полной неподвижностью: больной не мог не только повертывать туловища, но даже повертывать и выпрямлять голову, вследствие чего он производил впечатление человека, с упорством смотрящего на свои ноги. Достойно также внимания, что грудная клетка не принимала никакого участия в дыхании, которое совершалось исключительно благодаря Сокращениям грудобрюшной преграды. Чувствительность была значительно притуплена почти на всей поверхности тела, в особенности же в области шеи, спины и боковых частей груди. Мышечная сила рук была Заметно ослаблена. О субъективных ощущениях больной не мог давать показаний вследствие своего психического расстройства. Анамнеза получить не удалось. Больной умер от присоединившейся бугорчатки легких; вскрытия не было.
В другом случае были явления paralysis agitans и вместе с тем крайне резкая горбатость позвоночника в грудной и шей--ной областях, благодаря чему голова в сидячем и стоячем положениях представлялась опущенной прямо вниз, а взор устремлялся в пол. При этом замечалась поразительная неподвижность позвоночного столба: больной был лишен возможности сгибать и поворачивать туловище в стороны в сидячем или стоячем положениях, не мог поднимать головы и только слегка .двигал ею в стороны. По той же причине больной не мог без посторонней помощи повертываться в постели с одного бока на другой или переменить лежачее положение на сидячее; вставать же на ноги он мог, только помогая себе руками. В чувствительной сфере у больного наблюдались частые и разнообразные парестезии в области спины, груди и рук и ясное притупление чувствительности преимущественно на спине. Вместе с тем бросалось в глаза общее исхудание, в особенности же мышц лопаточных областей. Боли в позвонках не было ни при постукивании, ни при попытках сгибания. Дыхание было исключительно брюшным. Сила рук значительно ослаблена; да и в ногах заметна была паретическая слабость. Коленные рефлексы с обеих сторон были повышены. С течением времени паретическая слабость рук и ног значително усилилась, и развились
202
сгибательные сведения нижних, а затем и верхних конечностей. Впоследствии, как я слышал, больной умер от какого-то осложнения, причем вскрытие сделано не было. К сожалению, в этом случае причина, вызвавшая одеревенелость и искривление позвоночника, как и в предыдущем случае, осталась не выясненной; явления же paralysis agitans впервые начали обнаруживаться, как это часто случается с подобными больными, после внезапного и сильного душевного потрясения.
Из приведенных наблюдений ясно, что тяжесть болезни у моих больных была далеко не одинакова. Наименее тяжелую картину представляла первая больная, у которой заболевание в то же время было и более свежее по времени; последний же случай следует признать самым тяжелым. Тем не менее, не подлежит сомнению, что во всех приведенных случаях мы имели одно и тоже заболевание. В этом убеждает нас то, что во всех случаях картина и развитие болезни представляли много общего между собою. Так, почти во всех случаях существовали: 1) большая или меньшая неподвижность или, по крайней мере, недостаточная подвижность всего или только известной части позвоночного столба при отсутствии в нем резкой болезненности при постукивании и сгибании; 2) дугообразное искривление позвоночника кзади, преимущественно в верхней грудной области, причем голова представлялась выдвинутой вперед и опущенной; 3) паретическое состояние мышц туловища, шеи и конечностей, большею частью с небольшою атрофией спинных и лопаточных мышц; 4) небольшое притупление чувствительности преимущественно в области разветвления кожных ветвей спинных и нижних шейных нервов, а иногда и поясничных; наконец, 5) разнообразные признаки раздражения со стороны тех же нервов, в виде парестезии, даже местных гиперестезии и болей в спине и в шейной области, а также в конечностях и в позвоночном столбе; в последнем в особенности при долговременном сидении. В некоторых случаях были, очевидно, и признаки раздражения со стороны двигательных нервов. Так, в одном случае был род судорожного подергивания рук и ног; появлявшегося вслед за чувством щекотания в локтевых и подколенных областях; в другом случае имелись сведения верхних и нижних конечностей.
Что касается до этиологии болезни, то выдающимися причинами в тех трех случаях, в которых имеются анамнестические данные, были наследственность и травматическое повреждение спины. Последнее, очевидно, играло существенную роль в развитии болезни в 2 случаях; к сожалению, в третьем случае при собирании анамнеза на это обстоятельство не было обращено внимания больного, и потому остается неизвестным, не имела ли эта причина и в данном сучае известного значения? Наследственность отмечена во всех трех случаях. В дальней-
20J
шем обсуждении вышеприведенных наблюдений прежде всего возникает вопрос о причине искривления позвоночного столба. Так как об английской болезни здесь не может быть и речи, то уже из того обстоятельства, что во всех случаях было дугообразное, а не угловое искривление позвоночника и не было сколько-нибудь резкой местной болезненности его при давлении и постукивании, следует заключить, что мы имели дело с так называемым мышечным искривлением позвоночника. Мышечные формы искривления позвоночного столба, как мы знаем, в свою очередь, делятся по своему происхождению на привычные, ревматические и паралитические. О привычном искривлении позвоночника в моих случаях, конечно, не может быть и речи. Хотя некоторые больные и указывали, что им приходилось, много нагибаться при своих занятиях, но, без сомнения, правильнее всего смотреть на эти объяснения лишь как на желание больных подыскать наиболее доступную их пониманию причину для искривления позвоночного столба. Дело в том, что другие проявления болезни, как-то: боли, парестезии, анестезии, па-ретическая слабость, заметная, хотя и слабая атрофия мышц,, неподвижность позвоночного столба и совершенная невозможность произвольного его выпрямления положительно исключает мысль о привычном искривлении позвоночника. Надо думать поэтому, что указываемая некоторыми из больных необходимость частого сгибания спины, быть может, до некоторой степени только способствовала усилению искривления; основная же причина этого искривления во всяком случае должна быть иная. С другой стороны, в моих случаях не может быть речи и о ревматическом мышечном искривлении, которое обыкновенно развивается более или менее быстро, почти внезапно,, после простуды, одновременно с болями в спине, причем исправление кривизны позвоночного столба становится невозможным даже и на короткое время вследствие болей, появляющихся при малейшем сокращении произвольных мышц. Таким образом, путем исключения я прихожу к выводу, что в описанных выше случаях горбатость, по всей вероятности, обусловлена параличной слабостью мышц спины. В пользу этого, без сомнения, говорят: паретическая слабость в мышцах шеи, спины и конечностей (в особенности верхних), анестезия кожи и некоторая атрофия спинных мышц. Но в обыкновенных случаях паралитической горбатости имеются два существенно важные и характерные признака, отличающие это заболевание от всех других форм горбатости: отсутствие боли и неспособности выпрямлять позвоночник вследствие паралича мышц 1. В моих же случаях были налицо боли и парестезии в области спины,, шеи и отчасти в конечностях, а невозможность выпрямления позвоночного столба обусловливалась, повидимому, не столько-
1 Eulenburg, Real-Enzyklopadie. Bd. XI, ?. 5 ГА
204
•параличной слабостью мышц, сколько неподвижностью самого позвоночного столба. В самом деле, мои больные одинаково не могли ни выпрямлять, ни сгибать своего позвоночника в грудной области, так что при необходимости они нагибались всем туловищем, и притом всегда в несовершенной степени. Точно так же и боковые повороты позвоночника были резко ограничены или почти совсем невозможны. Достойно внимания, что, несмотря на значительную горбатость позвоночника кзади, не было заметного лордоза поясничной части или, по крайней мере, не было такого лордоза, как в обыкновенных случаях горбатости, зависящей от паралича спинных мышц х.
Очевидно, что в моих случаях была не простая параличная горбатость, т. е. не горбатость, обусловленная параличом спинных мышц, подобно описанной, например, Эйленбургом, а совершенно своеобразное заболевание, поразившее не только нервы шейных и спинных мышц и частью мышц конечностей, но также и самый остов позвоночного столба. Спрашивается, что же это за поражение?
Вопроса этого нельзя решить с положительностью до тех пор, пока случаи, подобные вышеприведенным, не будут исследованы и патологоанатомически. Тем не менее, руководясь клиническими данными, я желал бы высказать здесь свои соображения о патогенезе болезни, причем я хорошо сознаю, что соображения эти не могут иметь иного значения, как гипотезы, нуждающейся в патологоанатомической проверке.
Прежде всего необходимо иметь в виду, что ранними признаками болезни является чувство тяжести головы и невозможность поддерживать туловище в прямом направлении; поэтому уже в ранних периодах болезни развивается искривление позвоночника кзади. Рядом с этим очень рано же появляются и признаки раздражения со стороны шейных и спинных нервов, поражением которых, без сомнения, объясняются не только вышеуказанные паретические расстройства, с течением времени все более и более усиливающиеся, но и небольшая атрофия пораженных мышц. Но так как в то же время развивается и поразительная малоподвижность позвоночного столба, то, очевидно, что болезненный процесс, гнездясь в позвоночном канале, должен вовлекать в страдание одновременно как корешки, дающие начало спинным и шейным нервам, так и соединительнотканные части позвоночного канала.
Возможно, конечно, что с течением времени может присоединиться и поражение самого спинного мозга как последовательное заболевание. Этим, по всей вероятности, и объясняется бывшая в третьем случае местная болезненность позвоночника на уровне XI и XII грудных позвонков, паретическая слабость ног и усиление сухожильных коленных рефлексов.
См., например, известный случай Эйленбурга.
205
С вышеуказанным предположением о патологоанатомической основе поражения вполне согласуется и этиология страдания. Мы видели, что, кроме наследственности, предрасполагающей к заболеванию, травма спины играет роль важной непосредственной причины; и нельзя отрицать, что именно травма может сделать для нас легко понятным происхождение разлитого воспалительного процесса в клетчатке, окружающей твердую оболочку спинного мозга.
Что же касается до течения этого своеобразного болезненного процесса, то все больные, относительно которых имелись анамнестические сведения, единогласно свидетельствуют, что болезнь развивалась, хотя и медленно, но, повидимому, прогрессивно. Вероятно, в дальнейшем течении она оканчивается параличами и сведениями: по крайней мере, так можно заключить из третьего и пятого случаев, представлявших более развитую форму, нежели остальные.
Относительно лечения мои наблюдения, к сожалению, слишком неполны, чтобы можно было сделать какие-либо заключения. Могу лишь отметить благотворное влияние местных отвлекающих, в виде прижиганий прибором Паклена, на субъективные болезненные ощущения. Руководясь чисто теоретическими соображениями, можно было бы испытать и подвеши* вания, или вообще вытяжение позвоночника, но я, к сожалению, не имел возможности применить этот способ в моих случаях.
ОБ АНКИЛОЗИРУЮЩЕМ ВОСПАЛЕНИИ
ПОЗВОНОЧНИКА И БОЛЬШИХ СУСТАВОВ
КОНЕЧНОСТЕЙ
Несколько наблюдений особой формы заболевания, приводящей к одеревенелости позвоночного столба, опубликованных мною со времени 1892 г. в двух работах (см. «Врач», 1892, и «Neurologisches Zentralblatt», 1893, и «Deutsche Zeitschrift fur Nervenheilkunde», 1897), между прочим, побудили профессора Мари вместе с Асти х и профессора Штрюмпеля 2 заявить о своих наблюдениях, относящихся к поражениям позвоночного хребта с характером одеревенелости. Как ни приятно для меня сознание, что столь известные клиницисты по поводу моей статьи сделали со своей стороны публикацию своих наблюдений, я должен, однако, заметить, что наблюдения Штрюмпеля в этой области несколько отличаются от моих. Это обстоятельство отмечается и самим Штрюмпелем, вследствие чего он не решает окончательно Еопроса, вполне ли относятся его случаи к категории опубликованных мною наблюдений.
В общем страдание, о котором говорит проф. Штрюмпель, выражается процессом, постепенно и без болей приводящим к совершенному анкилозу позвоночного столба и бедренных суставов, так что голова, туловище и бедра плотно связываются друг с другом и совершенно одеревеневают, тогда как все другие суставы сохраняют свою нормальную подвижность. Само собою разумеется, что вследствие этого получается своеобразное изменение положения тела и походки. В наблюдении, приводимом Штрюмпелем в упомянутой статье, необходимо отметить, что и при глубоком хлороформном наркозе одеревенелость позвоночника не изменилась. Равным образом и движения в правом бедренном суставе оставались затрудненными, причем при вращении бедра чувствовалась теперь ясная крепитация. Достойно также внимания, что произведенные с терапевтической целью значительные пассивные движения не остались без вли-* яния на состояние больного. После наступившей первое время лихорадочной реакции и при лечении теплыми ваннами со-,
1 Marieet Astie, Presse med., Octobre 1898, № 89.
2 S t r u m p e 1 1 , Dtsch. Zschr. f. Nervenheilk., 1897.
20?
■стояние больного заметно улучшилось, и он уже мог свободнее ходить.
Существенным отличием этих случаев от моих наблюдений является то обстоятельство, что в последних совершенно не было поражения больших суставов и вместе с тем имелась ки-фотическая горбатость позвоночника в верхней грудной области и более или менее резко выраженные корешковые симптомы (боли, парестезии и пр.), тогда как в случае Штрюмпеля обращала на себя внимание, наоборот, поразительная прямота позвоночника, отсутствие резких корешковых симптомов и соучастие в поражении бедренных суставов. Штрюмпель, между прочим, выделяет свой случай от arthritis deformans позвоночника, описываемого Оппенгеймом, так как в этом случае дело идет о частном явлении общей формы заболевания и к тому же на первый план выступали боли и корешковые симптомы. Вслед за мною и Штрюмпелем были опубликованы подобные же наблюдения фон-Колером \ Веером 2, Беймлером 3, Н. Шаталовым 4, С. Поповым 6 и др.
Кроме того, в последнее время останавливается подробно на подобном же заболевании и Мари 6, описывая его в двух случаях под названием: «Spondylose rhizomelique». Еще ранее того (в сентябре 1897 г.) мне представилась возможность демонстрировать результаты патологоанатомического исследования в одном случае одеревенелости позвоночника7 . Последним сюда относящимся наблюдением, опубликованным уже после работы Мари, является случай Муттерера 8.
По взгляду Мари, характеристикой его «spondylose rhizomelique» является сопутствование полного сращения позвоночного столба более или менее ясно выраженным анкилозом суставов «корней конечности» при целости малых суставов конечностей.
В его случаях суставной анкилоз был вполне выражен собственно в бедренном суставе, который находился в несколько согнутом положении и обнаруживал полную одеревенелость. В плечевых же суставах обнаруживалось лишь затруднение движения. Спинной хребет был совершенно анкилозирован, причем в верхней его части существовало сильное кифотическое
1 Kohl er V., Charite-Annalen, 1887, v. XII.
2 Beer, Wiener med. Blatter, 1897, N 8 и 9.
3 В a u m 1 e r, Dtsch. Zschr. f. Nervenheilk., Bd. XII, Hft. 2.
4 Шаталов Н., Три случая так называемого натяжного анкило-зирующего воспаления позвоночника, Отчет о заседении Московского общества неврологов и психиатров, Обозрение психиатрии, 1898, № 9.
6 П о п о в С, Случай анкилоза позвоночника. Отчет о заседании Московского общества неврологов и психиатров, Обозрение психиатрии, 1899, № 1.
«Marie, Revue de medecine, 10 avril 1898.
7 Обозрение психиатрии, 1898, № 3.
β Mutterer, Dtsch. Zschr. f. Nervenheilk., Bd. XIV, Hft. 1и 2, стр. 144.
208
искривление, в то время как нижняя половина представлялась совершенно прямою. Небольшое уменьшение подвижности наблюдалось также в коленах. Кроме того, у больных существовали экзостозы в сакральной области и в телах шейных позвонков (пальпация через pharynx), уплощение грудной клетки и таза, неподвижность грудной клетки при дыхании и атрофии в спинных мышцах и в мышцах ягодиц.
Хотя описываемые Мари случаи и не вполне тождественны с вышеприведенными случаями, однако между ними и нет достаточно существенных различий для выделения их в совершенно самостоятельное заболевание. Повидимому, дело идет здесь о разновидности того самого заболевания, на которое указал Штрюмпель и которое, как я указал выше, отличается несущественными чертами от моих наблюдений, в которых не наблюдалось ни анкилоза больших сочленений, ни экзостозов позвоночника и в которых с резкостью выступали корешковые симптомы. Со своей стороны могу заметить, что я неоднократно имел случай наблюдать болезненное поражение с характеристическими признаками, указываемыми Штрюмпелем и Мари, которое, однако, я всегда отличал от того поражения, которое было описано мною под названием: «одеревенелости с искривлением позвоночного столба». Интереса ради я приведу здесь два таких наблюдения.
Больной Б., 34 лет, женатый, поступил в клинику 14 сентября 1896 г. Он сын чиновника, происходит из семьи, отличавшейся хорошим здоровьем, но отец его страдал запоем, хотя и дожил до глубокой старости. Сам больной всегда был здоров; на 21-м году жизни, т. е. в 1883 г., больной поступил на военную службу, где сначала служил писарем, а затем через 5 месяцев был переведен в Петербург в Артиллерийское управление. Через две недели по приезде в Петербург у него появилась впервые опухоль правого коленного сустава, вследствие чего он поступил в военный госпиталь, где пробыл около 8 месяцев. После этого, поправившись, ои снова поступил на службу. До 1886 г. по временам у него снова появлялась опухоль в том же суставе, исчезавшая под влиянием лечения. С этого времени и до настоящего заболевания больной вообще ничем не хворал, но иногда под влиянием простуды у него появлялась опухоль в коленных суставах, скоро, однако, исчезавшая. Сифилис отрицает. На 27-м году больной женился. Причину своего заболевания больной относит к простуде, так как ему подолгу приходилось заниматься счетоводством в плохо устроенном помещении, где он подвергался действию сквозного ветра. За два года до поступления в клинику он начал замечать боль в левом плечевом суставе, усиливавшуюся при поднимании и отведении руки. Вскоре появилась опухоль в том же суставе, которая распространилась быстро на шею и на правую руку. Больной лечился в Старой Руссе солеными ваннами, но особенного улучшения от этого лечения не было. За 3 месяца до поступления в клинику начала обнаруживаться слабость в нижних конечностях и болезненность в них при движении, вследствие чего при ходьбе он стал прибегать к палке.
При исследовании в клинике найдено следующее. Больной среднего роста, плохого питания. Костная и мышечная система развиты слабо. Общее исхудание тела. Замечается атрофия мышц в области шеи, верхних и нижних конечностей, причем в мышцах и тех, и других изредка удается заметить фибриллярные сокращения. В тазо-
14 В. М. Бехтерев
209
бедренных сустрвах обнаруживается почти совершенный анкилоз в полусогнутом положении, в плечевых суставах замечается значительная туго-подвижность. Неполные движения здесь возможны только вперед и немного в сторону, назад движения совершенно невозможны. При насильственном движении в плечевом суставе чувствуется резкая боль; равным образом при произвольных движениях верхними конечностями больной чувствует боль в плечевых суставах. Позвоночник почти совершенно неподвижен как в своей нижней части, так и в верхней; шея сгибается с трудом; туловище может лишь слегка нагнуться кпереди на счет поясничного его отдела. Положение туловища больного, благодаря анкилозу в тазобедренных суставах, согнутое вперед под тупым углом. Колени в сидячем положении слегка согнуты. Ходьба крайне затруднена вследствие малоподвижности и болезненности в суставах. Грудь плоская и почти не участвует в дыхательных экскурсиях, межлопаточный и кремастеровый рефлексы отсутствуют, брюшной и надчревный несколько усилены, равно как и подошвенный. Локтевые и коленные сухожильные рефлексы равномерно усилены. Мышечная реакция в атрофированных мышцах слегка ослаблена, но без следов извращения.
Из субъективных ощущений больной жалуется лишь на чувство стягивания в бедрах и болезненные ощущения в плечевых суставах. Следует отметить, что во время пребывания в клинике у больного без видимой причины появилась опухоль и болезненность, сначала в правом, а затем в левом голеностопном суставе, которая держалась в течение около 3—4 недель и затем постепенно исчезла. Спустя некоторое время появилась болезненность в тазобедренных и коленных суставах, причем правый коленный сустав начал обнаруживать припухлость. Со временем эти боли то опять затихали, то снова возобновлялись. Из терапевтических мер больному применялись соленые ванны, массаж и фарадизация пораженных конечностей; по временам при болях согревающие компрессы на суставах и смазывание их t-rae jodi, внутрь Natrium salicylicum и Kalium jodatum.
В конце концов под влиянием вышеозначенной терапии состояние больного заметно улучшилось, боли в суставах уменьшились и подвижность их увеличилась, так что к концу марта следующего года больной выписался из клиники в улучшенном состоянии.
Второй случай заключается в следующем.
Больной А., художник, 27 лет, холост, обратился ко мне за советом 17 мая 1898 г.
Отец умер 63 лет от апоплексии. Отец вел скромный образ жизни, был человек религиозный, алкоголизмом не страдал, сифилиса не имел. В отцовской линии не было ни нервных, ни душевных больных. Дед по матери был за несколько лет до смерти душевнобольным, умер от гангрены. Мать умерла на 52-м году от крупозного воспаления легких. Всех детей было 6; 2 умерли от случайных болезней в раннем возрасте и 4 живы. Две сестры замужние. Обе пользуются хорошим здоровьем, но старшая из них 5 лет тому назад страдала истерией. Младшая — незамужняя, здорова. Туберкулезных в родне не было. С детства наш больной был здоровым ребенком, зубы прорезались во-время и во-время больной начал ходить, золотухой не страдал и вообще тело было чистое; 8 лет перенес легкую форму натуральной оспы. 9 лет однажды по ошибке вместо воды проглотил цианистого и едкого кали в азотной кислоте (отец был фотограф по профессии). После отравления 13 часов находился в бессознательном состоянии, затем в течение недели были боли в животе. До 15 лет затем больной чувствовал себя здоровым. Сношений с женщинами больной никогда не имел и сифилисом не страдал. 15 лет начал чувствовать боли в коленных суставах, то в правом, то в левом. Через год эти боли приняли более острый характер, появилась водянка суставов; при дотрагивании чувствовалась болезненность. На 17-м году, через полгода после водянки, появились в мышцах судорожные
210
подергивания, затем появилась атрофия в левом бедре и голени. Коленные суставы были припухшими; опухоль белого цвета и тестоватая иа ощупь; при дотрагивании до опухоли появлялись рефлекторные подергивания. С появлением судорог, мешавших малейшему движению и сопровождавшихся резкой болезненностью, больной принужден был лечь в постель. Пролежал он 2 года. В эти два года левая нога резко атрофировалась. Благодаря массажу и ваннам страдание стало несколько регрессировать и спустя 2J/2 года от начала заболевания больной снова встал на ноги, но ходил на костылях. 3 месяца спустя после того, как больной стал ходить на костылях, заболевает без видимой причины и правая нога; в ией также стала обнаруживаться опухоль колейного сустава, появились судороги и боли молниеносного характера и очень резкое напряжение мышц бедра; по временам напряжение в бедре было таково, что больной не мог им сделать ни малейшего движения. Правая нога тоже в течение 2 лет не поддавалась никакому лечению; больной лежал в кровати почти неподвижно, так как всякое движение вызывало рефлекторно судороги в бедрах и болезненность по ходу п. ischiadici. Затем больной лечился на одесских грязевых курортах. Перед этим еще он стал ходить на костылях. После двухмесячного грязевого лечения судорожные подергивания и боли уменьшились, движение увеличилось, опухоли рассосались. После грязевых ванн было назначено купанье в море, но после 18-го купанья дело приняло очень скверный оборот: вдруг появилось напряжение в мышцах обоих бедер и сильная болезненная рефлекторная возбудимость; больной слег в постель; до ног его нельзя было дотронуться. В таком положении он вернулся в Петербург. Здесь по назначении ванн и иода состояние несколько улучшилось, но все же больной снова пролежал в кровати около 2 лет. Когда больной встал на костыли, то, как он говорит, «при освидетельствовании нижний отдел позвоночника оказался неподвижным». Тазобедренные суставы почти стали неподвижными; затем после применения гимнастики движение тазобедренных суставов несколько улучшилось. В 1894 г. больной был в Саках, а потом поселился в Крыму. Все эти три года обострений не было. Каждое лето больной был в Саках. За три года состояние улучшилось.
Советовался с Вельяминовым, Склифосовским, Мультановским, Шер-шевским, Ратимовым, Субботиным, Захарьиным, Дьяконовым, Мочут-ковским, Поповым, Павловым, Гопадзи и другими. Одни из них, как Вельяминов и Домбровский, предполагали страдание оболочек спинного мозга, тогда как большинство других признавали воспаление суставных сумок ревматического характера.
Все время заболевания температура была не выше 37,1° и 37,2°, во время припадков понижалась до 36° с десятыми, иногда даже до 35,8°. Желудок работал хорошо, анализы мочи не давали никаких особых патологических данных. Мочеотделение все время было нормально.
Status praesens. При осмотре больного левая нижняя конечность кажется несколько короче правой. Когда больной стоит, то пятка, левой ноги не совсем касается пола. Вместе с тем левая конечность кажется несколько худее правой. Точное измерение окружностей бедер и голеней дало следующие цифры:
Правое бедро .... 51 см, левое......50 см
Правая голень .... 34,5 » левая......34 »
Правое плечо .... 32,5 » левое......32 »
Правое предплечье . 26 » левое......26 »
Измеряя длину правой и левой конечности, можно убедиться, что левая нога короче правой на 0,5—1 см. Голеностопные суставы оба нормальны и все движения в них возможны. Движения в коленных суставах ограничены, именно ограничено несколько сгибание; согнуть колени больной может только до прямого угла, разгибание свободно. В обоих тазобедренных суставах движения резко ограничены: ни сгибание, ни разгибание
14* 21.1
невозможно. Приведение и отведение бедер совершаются в самых незначительных размерах. Движения позвоночника возможны только в шейной части и небольшое движение в верхне-грудной области, остальная часть позвоночника совершенно неподвижна, больной не может нагнуться вперед, назад, а равно отсутствуют и боковые, и вращательные движения позвоночника. При этом позвоночник представляется равномерно изогнутым кзади; при постукивании безболезнен. Стоит больной, слегка нагнувшись вперед, с полусогнутыми коленями. При ходьбе позвоночник неподвижен, движения таза незаметны, тазобедренные суставы как бы не функционируют и движения ног происходят только в коленных и голеностопных суставах; при этом, ступая на левую стопу, больной становится более на носок, пятка же совсем пола не касается. Если больного, когда он стоит, попросить сесть, то, садясь, он держится руками за ручки кресла и таким образом опускается в кресло, причем таз, поясница и нижне-грудная часть позвоночника остаются совершенно неподвижными; вместе с тем больной не может и встать иначе, как поднимаясь с кресла на руках. По словам больного, его позвоночник к постукиванию не был болезнен во все время заболевания. Нервные стволы к давлению тоже не болезненны, при этом больной заявляет, что ηπ. ischiadici бывают болезненны к давлению лишь во время обострения процесса; то же делается и с бедренным нервом.
Мышечная сила по динамометру правой руки 120 — 100 фунтов, левой 80 — 90 фунтов. Мышечная сила нижних конечностей довольно хороша. Исследование чувствительности тактильной, болевой и температурной показало, что все три вида чувствительности вполне нормальны. Мышечное чувство тоже не представляет уклонений от нормы. Электрическая возбудимость нервов и мышц не дала особенных данных, фарадическая возбудимость мышц левого бедра и голени несколько ниже, чем на правой стороне, гальваническая одинакова и равна 1,5— 2 тА; при этом при замыкании катода сокращение мышц повсюду получается при меньшей силе тока, чем замыкание анода (КЗС>АЗС). Мышцы бедер напряжены. Кожные рефлексы: подошвенный левый выражен резко, правый слабее, такое же отношение представляет рефлекс cremaster; ягодичный рефлекс выражен слабо вследствие напряжения ягодиц. Брюшной и надчревный рефлексы выражены слабо, подреберья втянуты и мышцы живота сокращены. Лопаточного рефлекса нет.
Сухожильные рефлексы верхних конечностей нормальны, коленные рефлексы умеренно повышены, слева сильнее, чем справа, рефлекс с ахиллова сухожилия с левой стороны получается сильнее, чем с правой; клонуса коленной чашки, а равно и клонуса стоп вызвать не удается.
Больной заявляет, что он может ходить в настоящее время около 5 часов в сутки, причем если он ходит меньшее время,то чувствуется в ногах напряжение, если большее и устанет, то в ногах на другой день появляются боли. Стоять долго не может тоже вследствие болей. Органы чувств нормальны. Мочеотделение правильно, равно как и отправления кишечника. Половых сношений не имел, но эрекции и поллюции наблюдаются в месяц 1—2 раза.
Оба случая, из которых наблюдение над первым относится еще к 1896 г., в существенных частях повторяют почти все характеристические особенности наблюдений, представленных Мари и Штрюмпелем; но в то же время они представляют резкое отличие от формы болезни, описанной мною под названием одеревенелости позвоночного столба с его искривлением в том отношении, что здесь выступает участие в поражении больших сочленений. В моих случаях этой формы заболевания дело даже начало развиваться с поражения сочленений, предшествовавшего еще задолго до развития самой болезни и временно
212
то обострявшегося, то снова затихавшего, пока не развилось постоянное поражение бедренных суставов, перешедшее затем и на позвоночник. Судя потому, что при рассматриваемой форме дело доходит до анкилоза суставов, необходимо признать, что сущность заключается в оссифицирующем процессе, поражающем суставные поверхности больших сочленений и позвонков. Взгляд этот получает подтверждение, между прочим, в том обстоятельстве, что в случае Мари костное сращение бедренного сустава было констатировано при резекции этого сустава. Кроме того, в наблюдениях того же Мари отмечается присутствие экзостозов в крестцовой области и в телах шейных позвонков.
Надо, впрочем, заметить, что суставной процесс иногда вовлекает в поражение и соседние мягкие части, благодаря чему. развивается своеобразная ригидность мышц, соседних с пораженными суставами, и как последствие поражения сустава появляется атрофия мышц х.
Далее, в отличие от случаев описанной мною одеревенелости позвоночника, в которых первенствующую роль в этиологии играет наследственность, травма и, может быть, сифилис, должно заметить, что в рассматриваемой форме заболевания очевидную роль играют так называемые ревматические причины. Это ясно, между прочим, выразилось в случае Мутте-рера, в котором больной кузнец обвинял в происхождении своей болезни необходимость частых переходов из жаркой мастерской в холодный фабричный двор. В одном из моих случаев также больной приписывает происхождение своей болезни постоянным сквознякам, которым он подвергался в помещении, где он должен был подолгу сидеть, занимаясь счетоводством. С другой стороны, влияние наследственности, травма и сифилис в обоих случаях отсутствовали.
Наконец, и течение болезни в настоящем случае представляет особенности, не свойственные описанной мной одеревенелости позвоночника. В то время как в этих случаях анкилоза позвоночника и больших сочленений болезнь постепенно распространяется по позвоночнику снизу вверх, в описанных мною случаях одеревенелости обыкновенно раньше и сильнее всего поражалась верхне-грудная область позвоночника.
Упомянем также, что при описываемом заболевании, по-видимому, не обязательно кифотическое искривление позвоночника, как показывает по крайней мере случай Штрюмпеля; кроме того, и корешковые симптомы здесь обычно не представляются резко выраженными.
Итак, присутствие оссифицирующего поражения больших сочленений, восходящее распространение болезненного процесса, ревматоидные условия происхождения болезни, непостоянство
1 Беер (Wiener med. Blatter, 1897, № 8 и 9) придает даже главное значение поражению мягких частей, с чем, однако, нельзя согласиться.
213
кифоза, нерезкое выступление или даже отсутствие корешковых симптомов, не говоря о многих других частностях, отличают настоящее заболевание от описанной мною формы под названием «одеревенелости позвоночника с его искривлением».
В заключение нельзя не отметить, что при рассматриваемом заболевании поражаются не только те суставы, которые находятся в корнях конечностей, но и другие, как, например, коленный и голеностопный. В обоих наших случаях болезнь даже начала развиваться с поражения коленных суставов, а в первом случае поражение голеностопного и коленного суставов с болями и припухлостью обнаружилось даже в течение полного развития болезни. Отсюда ясно, что название spon-dylose rhizomeli que, данное этой болезни Мари, не совсем отвечает действительности; равным образом и название Штрюм-пеля неудовлетворительно в том отношении, что указывает на ограничение болезни бедренными суставами и позвоночником, вследствие чего я предпочитаю ему название хронического анкилозирующего воспаления больших суставов и позвоночного столба.
О ЗВУКОВОЙ ПРОВОДИМОСТИ ПРИ ПОСТУКИВАНИИ
КОСТЕЙ ЧЕРЕПА И ПОЗВОНОЧНИКА КАК МЕТОДЕ
ИССЛЕДОВАНИЯ НЕРВНЫХ БОЛЕЗНЕЙ
В последнее время были опубликованы две работы, относящиеся к исследованию звуковой проводимости черепных костей, — д-ра В. Н. Окунева и Г. Н. Габричевского. Первый автор х применял исследование звуковой проводимости главным образом для определения гнойного воспаления сосцевидного отростка височной кости при страданиях среднего уха. Для этой цели он пользуется обыкновенной отоскопической трубкой, наконечник которой он вставляет себе в ухо, каучуковую же воронку ставит на сосцевидный отросток височной кости. Затем, ударив камертоном по какому-либо предмету, он ставит его или на темя или немного впереди и ниже теменного бугра. При исследовании на месте пораженного сосцевидного отростка звук камертона через отоскопическую трубку, по автору, слышится всегда менее ясно, нежели на месте здорового сосцевидного отростка.
Все результаты своего исследования автор сводит к следующим главным положениям:
1. При решении вопроса о трепанации сосцевидного отростка височной кости всегда следует испытывать и костную проводимость.
2. Изменение силы звука камертона при передаче через кости в смысле ослабления или заглушения ее должно считаться несомненным признаком глубокого страдания кости (гнойное гнездо, костоеда, новообразование и т. д.).
3. Такие же изменения костной звукопроводимости должны наблюдаться, по всей вероятности, и при глубоких страданиях всех остальных костей человеческого тела, но, по неимению подходящего материала, мне еще не удалось проверить это предположение.
4. Указываемый мною признак изменения костной звукопроводимости позволяет трепанировать раньше, особенно в сомнительных случаях с вялым, хроническим течением болезни.
1 Врач, 1893, № 48 и 49.
215
Поэтому он должен способствовать более, благоприятному исходу после трепанаций, так как позволяет определять страдания костей еще тогда, когда в них нет видимых изменений.
Д-р Габричевский 1 исследовал звукопроводимость черепа с помощью особого аппарата «пневматоскопа», предназначенного первоначально для исследования звуковых колебаний воздуха, заключенного в дыхательных путях. Воронка этого прибора приставляется к открытому рту и звуковые волны, получаемые с поверхности грудной коробки и черепа или обыкновенным постукиванием, или приставлением звучащего камертона на грудь или череп, выслушиваются исследователем с помощью двух резиновых трубок, сообщающихся с воронкой прибора. Еще в 1890 г. в Берлинском медицинском обществе автор высказал мысль, что с помощью его прибора можно исследовать не только грудь, но и голову. Вскоре затем автор на практике убедился в возможности распознавать с помощью своего метода гнойное воспаление сосцевидного отростка височной кости, обусловливающее слабое звучание камертона. Впоследствии автор мог убедиться в распознавательном значении своего метода и в других случаях, между прочим и при мозговых опухолях. Д-р Габричевский в своей статье вскользь замечает, что при исследовании звукопроводимости черепных костей он нередко пользовался обыкновенным двуушным американским стетоскопом, который приставлялся им к средней линии головы, причем сравнивалось звучание камертона на симметричных частях черепа. Автор отдает преимущество своему методу, так как звучание камертона гораздо слышнее при пнев-матоскопическом исследовании, чем с помощью стетоскопа. При первом способе, однако, получаются дыхательные шумы, которые надо исключать или путем навыка со стороны врача, или же задержкой дыхания со стороны больного на время исследования.
В заключение своей статьи автор заявляет, что 1) способ (пневматоскопический и стетоскопический) исследования звуковой проводимости черепных костей и костей вообще был выработан впервые им и что 2) с помощью этого способа ему первому удалось показать возможность распознавания определенных патологических изменений мозга и черепа.
Оставляя совершенно в стороне вопрос о приоритете как менее всего интересный для читателей, я со своей стороны должен заметить, что исследование звукопроводимости черепных костей заслуживает большого внимания в нервной патологии. Я думаю даже, что вряд ли найдется какой-либо невропатолог, у которого не проскользнула когда-либо мысль относительно желательности непосредственного выслушивания головы при черепных заболеваниях.
1 Врач, 1893, № 52.
216
Что касается меня, то уже почти с первых шагов своей врачебной деятельности, следовательно, задолго до вышеприведенных исследований, я пытался применить метод выслушивания к 'голове и уже давно пользуюсь для этой цели обыкновенным стетоскопом, который я приставляю к средней части темени; самое же постукивание произвожу пальцем по тем частям головы, которые желаю исследовать, причем выслушиванием сравнивается звукопроводимость симметричных или соседних частей головы.Там, где нужно исследовать самое темя, я отодвигаю стетоскоп ближе кпереди и ставлю его несколько выше лба, после чего выслушиваю звук, получаемый при постукивании пальцем по теменным областям головы. При этом методе мне удавалось неоднократно убедиться в изменении звуковой проводимости не только в тех случаях, где дело касалось поражения самой кости, но и в тех случаях, где можно было на основании клинических данных локализировать место мозгового поражения. При этом звук, получаемый при постукивании областей, соответствующих поражению, представлялся всегда заметно притуплённым, менее громким и во всяком случае не отдавался так в ухо, как звук, получаемый при постукивании здоровых частей головы.
Сейчас я наблюдаю случай с гнездным поражением мозжечка, где таким способом удается обнаружить ясное притупление звука в задней части головы, в ближайшем соседстве с затылочным бугром. В другом случае меньеровой болезни обнаруживается подобное же притупление на сосцевидном отростке пораженной стороны.
Я не привожу здесь других случаев, бывших под моим наблюдением, так как не считаю свои наблюдения законченными. Но я бы очень рекомендовал лицам, желающим посвятить свой труд исследованию звуковой проводимости черепа, не забывать и столь простого приема исследования с отоскопической трубкой, доступного решительно всякому практическому врачу, которому более сложный аппарат составляет всегда, некоторое бремя в практической деятельности. Здесь же добавляю, что с неменьшим удобством можно пользоваться методом выслушивания и в ином направлении, а именно: стетоскоп может быть помещаем на исследуемых местах черепа, постукивание же пальцем в таком случае следует производить по одному из удаленных участков черепа по направлению передне-задней линии; в этом случае звук на месте поражения слышится заметно слабее, нежели на соседних местах, а равно и на симметричных местах здоровой стороны.
Что касается камертонного исследования звукопроводимости черепных костей, то нельзя не признать, что этот метод дает в руки врача очень удобное, средство выслушивания звукопроводимости черепных костей. Но так как при этом выслушивается передаточный звук камертона, то я придаю значение не столько
217
тем аппаратам, с помощью которых выслушивается этот звук, сколько тому, чтобы самый звук камертона отличался необходимым постоянством. В этом отношении точность исследования очень мало гарантируется приставлением к черепу обыкновенного камертона, который заставляют • звучать предварительным ударом о тот или другой предмет. С целью устранения возможных в этом отношении неточностей я озаботился устройством постоянно звучащего камертона, который в то же время весьма удобно и легко приставляется к костям черепа. Модель такого камертона недавно по моей мысли была приготовлена д-рами Жуковым и Добротворским и затем выполнена
в окончательном виде в электротехнической мастерской Подобедова (рис. 1).
Прибор представляет собой не что иное, как обыкновенный камертон (а), при одной вилке которого, на особом деревянном штативе (Ь), укреплена катушка с электромагнитом. Ради удобства приставления к черепу такого камертона к деревянному штативу его приделана ручка (с), за которую и держит наблюдатель, исследующий звукопроводимость черепа.
Всякому понятно, что, соединив катушку электромагнита с гальванической батареей, мы заставим камертон звучать непрерывно и равномерно, а это и нужно для исследования звуковой проводимости головы. Поставив такой звучащий камертон на теменную область головы, мы можем обыкновенным или двойным стетоскопом (что, на мой взгляд, несущественно) обойти всю голову, выслушивая ее различные области. В этом случае распознавательным признаком поражения черепных костей и подлежащих областей мозга является белее или менее явственное заглушение тона, что выясняется путем сравнения звукопроводимости симметричных и соседних областей черепа. Между прочим, в вышеупомянутых случаях с поражением мозжечка и с меньеровой болезнью на месте притупления костного звука обнаруживалось вполне явственное заглушение тона. Подобное же заглушение тона на месте сосцевидного отростка я нахожу и в одном из наблюдаемых мною случаев с новообразованием, развившимся в глубине височной кости.
Должно при этом заметить, что оба вышеуказанных метода применимы к исследованию не только костей черепного свода, но и к исследованию позвоночного столба.
Рис.
218
В месте с выслушиванием звукопроводимости головы в своей клинической практике я обычно пользуюсь и методом постукивания для выяснения местной болезненности головы и позвоночника.
Как известно, уже давно местная болезненность черепа и костей позвоночника принималась во внимание при постановке местного диагноза черепно- и спинномозговых заболеваний, но, несмотря на то, до настоящего времени в этом отношении произведено весьма мало исследований.
Сколько мне известно, на немецком языке несколько лет тому назад появилась специальная работа относительно значения местной болезненности головы, сделанная под руководством профессора Э. Менделя; в отечественной же литературе я не могу припомнить себе ни одного исследования, специально посвященного этому предмету. Между тем не подлежит сомнению, что в указанном отношении существует еще много невыясненного и неустановленного, в силу чего я и считаю нелишним сообщить здесь вкратце результаты своих наблюдений по этому предмету.
На мой взгляд прежде всего заслуживает внимания самый способ постукивания черепа и позвоночного столба. В этом отношении я различаю поверхностную и глубокую перкуссию. В первом случае производится слабое постукивание врачебным молотком или пальцем непосредственно по позвоночнику, тогда как во втором случае постукивание производится с большой силою и обязательно должно производиться через плессиметр.
Постукивание первого рода открывает нам болезненные процессы, гнездящиеся преимущественно в самых позвонках (например, местный туберкулез, периостит, сифилитический склероз кости и т. п.); ввиду этого самое постукивание должно быть производимо с большею осторожностью, так как нередко бывает уже достаточно не только слабого постукивания, но даже некоторого надавливания пальцем на кости черепа или на позвонки, чтобы вызвать у больного резкую боль в месте давления.
Только что указанный способ исследования, а равно и диагностическое его значение, конечно, достаточно хорошо известны невропатологам и потому нет надобности здесь распространяться по этому предмету. Иначе дело обстоит с глубокой перкуссией черепа и позвоночного столба, которая, сколько мне известно, мало или вовсе не практикуется для выяснения местного диагноза черепно- и спинномозговых поражений. Между тем значение ее в только что указанном отношении, как я убедился на основании личных наблюдений, неоспоримо.
Само собой разумеется, что глубокая перкуссия неприменима там, где по причине поражения самого черепа или позво-
219
ночника имеется резкая болезненность их ко всякому, даже незначительному постукиванию; во всех же других случаях глубокая перкуссия может быть применима совершенно безнаказанно и с большой пользою. При этой перкуссии, благодаря сотрясению значительной части черепа или позвоночного столба с их содержимым, нередко является возможность открыть болезненность там, где обыкновенным способом поверхностного постукивания не удается обнаружить даже и малейших следов болезненности. Так, мне неоднократно удавалось этим путем открывать у эпилептиков болезненность той или другой теменной области, точно так же при поражении задних частей мозга мне удавалось открывать болезненность затылочной области головы и т. д.
Не менее полезным метод глубокой перкуссии оказывается при тех или других спинномозговых поражениях.
Как известно, при хронических поражениях спинного мозга с местным характером, не затрагивающих самый позвоночный столб, обыкновенное, т. е. сравнительно слабое, постукивание по позвоночнику в огромном большинстве случаев не открывает в нем никакой чувствительности или болезненности. При более же сильном постукивании через плессиметр удается и в этих случаях открыть чувствительную область в той или иной части позвоночника, причем я неоднократно убеждался в одних случаях на основании сопровождающих клинических явлений, в других —на основании результатов последующего вскрытия, что чувствительная или болезненная область в общем соответствовала месту поражения спинного мозга. Таким путем мне удавалось во многих случаях открывать более или менее точно место застаревших сифилитических процессов в спинном мозгу и его оболочках, место старого миэлитического процесса и т. п.
Диагностическое значение глубокой перкуссии выясняется в особенности в тех случаях, где по другим проявлениям болезни мы не имеем возможности судить даже с приблизительной точностью о месте спинномозгового поражения.
Как известно, важнейшим указателем на место поражения спинного мозга, кроме распределения расстройств чувствительности и двигательного паралича, служат результаты исследования сухожильных и кожных рефлексов; но первые легко и с особенным постоянством вызываются только с сухожилий мышц нижних конечностей, на верхних же конечностях они получаются у здоровых менее постоянно; что же касается кожных рефлексов, то в нормальном состоянии многие из них нередко совершенно отсутствуют или по крайней мере ослаблены в такой степени, что исследование их не может дать никаких более или менее точных указаний относительно места поражения спинного мозга. В этих-то случаях, определяя местную чувствительность или болезненность позвоночного столба с помощью
220
глубокой перкуссии (через плессиметр), мы имеем возможность открыть локализацию спинномозгового поражения даже при том условии, если для решения вышеуказанной задачи мы не можем руководствоваться какими-либо другими данными клинического исследования.
Мне, разумеется, нет надобности пояснять здесь, что более точное определение места спинномозгового поражения имеет не один лишь теоретический интерес, но и чисто практический, так как вместе с установлением локализации спинномозгового поражения является возможность применения, кроме общей, еще и местной терапии того или иного характера.
О ЗНАЧЕНИИ СОВМЕСТНОГО УПОТРЕБЛЕНИЯ БРОМИДОВ И ADONIS VERNALIS ПРИ ПАДУЧЕЙ
При падучей, как известно, применяется целый арсенал средств, оказывающих в известных случаях благотворное действие. Из всех этих средств заслуженною славою пользуются бромистые препараты, которые, будучи применяемы в больших дозах, во многих случаях оказывают, несомненно, благодетельное влияние на течение эпилепсии. Должно, однако, заметить, что лишь в сравнительно небольшом числе случаев при одном лечении бромидами припадки эпилепсии исчезают совершенно; в значительно большем числе случаев они лишь ослабляются в интенсивности и частоте, в других же случаях лечение бромидами вовсе не оказывает благотворного результата на течение эпилепсии, а в исключительных случаях, чему примеры я встречал неоднократно, даже оказывается прямо вредным, усиливая и учащая приступы. Если даже не принимать в соображение последние случаи, как случаи исключительные, то все же прославляемое лечение эпилепсии бромидами далеко не составляет верной панацеи от этого тяжелого недуга. Так как другие средства (атропин и пр.), применяемые при эпилепсии, обладают еще менее верным действием, то не без основания за последнее время стали обращаться к совместному применению бромидов с другими средствами. Так, В. Пуле г предложил лечение эпилепсии совместным употреблением бромидов с калабарским бобом или сернокислым эзе-рином, с пикротоксином, с белладонной или сернокислым атропином и с наперстянкой, причем последнее средство автором применяется в случаях, когда дело идет о «сердечной эпилепсии» с одновременным повышением кровяного давления. Поэтому автору, каждое из упомянутых средств в отдельности приносит гораздо менее пользы, чем какое-либо из них в соединении с бромидами, равно как и последние без какого-либо из упомянутых средств. В тяжелых случаях эпилепсии автор советует прямо начинать с одной из этих комбинаций. Что же касается выбора той или другой комбинации в каждом данном случае, то в этом отношении лишь отчасти можно руководиться состоя-
1 Р о и 1 е t V., Bull. gen. de therap., 1891, № 10; Новости терапии, 1891, стр. 249.
222
нием зрачков, т. е. их расширением или же сужением, в большинстве же случаев приходится действовать ощупью, на что требуется в крайнем случае не свыше 3 месяцев. Между прочим, как преимущество такой комбинации бромидов с выше названными средствами автор выставляет то обстоятельство, что благодаря ей можно значительно уменьшить дозу бромидов. В доказательство действительности выше названного способа лечения автор приводит 5 случаев эпилепсии с благоприятным результатом такого лечения.
Затем в позднейшее время, а именно в апреле прошедшего (1893) года, проф. Пауль Флексиг х опубликовал метод лечения эпилепсии, состоящий из комбинации лечения опием и лечения бромидами. В этом случае начинают с лечения опием, который назначается первоначально в малых приемах (порошка или экстракта 2—3 х 0,05 pro die), и постепенно возвышает до 1,0 pro die и даже еще выше в дозах от 0,25 до 0,35. После приблизительно 6 недель опий внезапно отменяется и тотчас же взамен того назначают бром в больших дозах (7,5 pro die). После продолжительного применения больших доз бромистых препаратов (около 2 месяцев) приемы последних постепенно уменьшаются до 2,0 pro die. Автор придает особое значение внезапной смене опия бромом. Обыкновенно припадки прекращаются со времени замены опия бромом, хотя в некоторых случаях они исчезают или ослабевают еще во время лечения опием. Вообще же лечение опием, повидимому, является, подготовительным в том смысле, что оно облегчает действие брома, т. е. делает его более интенсивным. По автору, этот метод в 2 случаях застарелой эпилепсии приостановил приступы совершенно и на продолжительное время, в других же случаях, в которых по другим основаниям лечение опием не могло быть достаточно продолжительным, был ясный, хотя только преходящий успех. В заключение своей статьи автор приводит один случай старой и упорной эпилепсии, излеченный по вышеуказанному методу.
Со своей стороны я должен заявить, что уже в течение многих лет мне удавалось применять с большим успехом при эпилепсии смесь бромидов с Adonis vernalis. К этой комбинации я пришел на основании того соображения, что в основе припадков эпилепсии должны лежать вазомоторные изменения в черепном мозгу с характером активной гиперемии черепной полости 2, тогда как действие бромидов при эпилепсии, повидимому, сводится главным образом к понижению возбудимости мозговых центров и, в частности, мозговой коры. Ввиду этого мне и казалось наиболее целесообразным соединить действие бромидов с таким средством, которое, повышая кровяное давление, сжимает до некоторой
х F1 ее h s i g P., Neurol. Zbl., 1893 (Orig. Mitt.).
2 См. мою статью на основании произведенных в моей лаборатории исследований д-ра А. Ф. Тодорского в № 22 Neurolog. Zbl., 1891, и статью д-ра Тодорского в «Медицинском обозрении» за 1894 г.
223
степени просвет самих сосудов, а таковым является, между прочим, Adonis vernalis.
Доза последнего в моих случаях равнялась от β 3 ДО oj при sjj—ajjj бромидов на gvj воды с таким расчетом, чтобы микстура принималась от 4 до 6 раз, а в иных случаях и до 8 раз в день. Как sedativum я присоединяю к этой микстуре нередко кодеин в количестве gr. jj — jjj на з-vj микстуры. В таком размере мои больные переносили лечение без всяких дурных последствий в течение многих месяцев беспрерывно.
На основании наблюдения многих случаев эпилепсии я могу сказать, что вышеуказанное лечение нередко почти сразу прерывает приступы эпилепсии, в других случаях оно резко уменьшает число и силу приступов. За много лет своей практики я не видел собственно ни одного случая эпилепсии, где бы систематически проведенное лечение вышеназванной смесью не оказало своего благотворного влияния в том или ином отношении.
В доказательство сказанного я позволю себе привести два следующих наблюдения, взятых ради примера.
Больной Г., 17 лет, наследственно предрасположенный со стороны отца, перенесшего психоз. Будучи совершенно здоровым мальчиком, на 9-м году жизни он упал с гимнастики, ударившись затылком о стену. Ушиб, повидимому, прошел бесследно, но на 15-м году жизни в марте 1892 г. с ним неожиданно происходит во сне судорожный приступ эпилепсии. В том же году впервые домашние заметили ненормальное возвышение кости на темени, которое, по словам больного, он замечал у себя и за несколько времени перед тем. В апреле развился второй приступ и вслед за тем припадки стали повторяться сначала довольно редко, а со временем, в особенности в течение 1893 г., они резко участились. Припадки большею частью случались во сне, хотя бывали и днем, причем вспоследствии они стали сопровождаться непроизвольным мочеиспусканием. При исследовании в ноябре 1893 г. найдено правильное сложение организма и отсутствие каких-либо изменений со стороны внутренних органов, зрачки слегка неравномерны; на темени обнаруживается довольно значительное разлитое местное возвышение кости, не болезненное к давлению и постукиванию, расположенное большею своею частью справа от стреловидного шва. Впрочем, при повторном постукивании удается иногда обнаружить местную болезненность в задней части опухоли. Надобно заметить, что родители больного обратились ко мне за советом по поводу предложенной им операции трепанирования черепа в теменной области, после того как систематическое лечение бромидами и другими средствами не оказало никакого влияния на течение болезни.
Осмотрев больного, я сказал родным, что с операцией можно еще и повременить, так как предварительно следовало бы испытать, кроме применявшихся средств, еще и другие способы лечения. Мать неохотно согласилась на новую «проволочку» времени, тем не менее не отклонила сделанного предложения. Больному были предварительно назначены для испытания большие дозы бромидов с йодистым потассием, но они не оказали никакого влияния на ход болезни, так что мать уже отказывалась продолжать лечение. Ввиду этого вместо прежней микстуры больному было назначено Inf. Adonis vernalise бромидами. С трудом можно было уговорить мать больного продолжать фармацевтическое лечение, от которого она пока не видела никаких результатов и потому настаивала на операции. С назначением Inf. Adonis vernalis с бромидами дело, однако, быстро изменилось к лучшему и припадки вскоре совершенно прекратились. Обозре-
224
ниё припадков в хронологическом порядке лучше всего пояснит сказанное:
1892 г. Лечение бромидами и другими средствами. 28 марта 1 судорожный припадок во сне.
12 октября замечена крайняя слабость, какая бывает после припадка, который мог быть просмотрен.
30 октября 1 судорожный припадок во время игры на рояле.
4 декабря 1 судорожный припадок на катке. Целый день после того бред.
1893 г.
10 января 2 припадка.
28 » 1 припадок во сне.
6 апреля 1 припадок во сне.
29 » 4 припадка вечером. 15 июня 1 припадок во сне. 19 » 1 припадок во сне.
7 июля 1 припадок во сне.
2 сентября 1 припадок во сне.
22 » 1 припадок во сне. 2 октября 1 припадок во сне.
Назначено мною лечение большими дозами бромидов с йодистым по-тассием.
19 октября 1 припадок во сне.
1 ноября 1 припадок во сне.
6 » 1 припадок во сне.
Лечение Inf. Adonis vernalis 3s — §vj, Potassii bromati 3JJS. Potas-sii jodati gjs — no 4 раза вдень.
4 декабря 1 припадок во сне очень слабый.
23 » тот же раствор увеличен до 6 приемов в день. 1 припадок во сне также очень слабый.
22 января Кратковременный туман в голове с легкими судорогами в горле, но без потери сознания. Затем припадки прекратились совершенно.
Последний раз я видел больного 24 мая. У него попрежнему опухоль на темени, зрачки слегка неравномерны, но самочувствие превосходное и припадков не обнаруживается вовсе. Он уже начал умственно заниматься, тогда как прежде заниматься совершенно не мог, вследствие вызываемого этим обострения припадков.
Больной Ш., 25 лет, из Нижнего Новгорода, происходит от нервной матери. Припадки эпилепсии с 10-месячного возраста. Отчего они произошли, больной сказать не может. Припадки обыкновенно начинаются чувством безотчетного страха и сердцебиением. Иногда припадок этим и ограничивается. В таком случае больной, почувствовав внезапный испуг, прижимается к матери и затем через 2 минуты совершенно освобождается от приступа, сохраняя сознание о происшедшем. В других случаях припадки сопровождались потерей сознания и судорогами с прикусом языка. Кроме того, время от времени с больным случались судорожные приступы в течение ночи, сопровождавшиеся сначала чмоканием, а затем общими судорогами с пеной у рта и прикусыванием языка. Приступы малой эпилепсии с больным случались обыкновенно по нескольку раз в день. Судорожные же приступы с больным случались раза по 2, по 3 в месяц. После приступа больной быстро оправляется, не чувствуя большею частью и головной боли. Объективно замечается лишь некоторая неравномерность зрачков. В июле 1890 г. с больным случился без видимой причины сильный приступ, во время которого больной оставался в бесчувственном состоянии с полчаса. С этого времени больной обратился ко мне в Казань за советом, причем ему были назначены бромистые препараты в больших приемах, ванны в 28° R и души —от 27° до 25°R. После того приступы заметно ослабели в силе и стали реже, но тем не менее продолжались еще довольно часто. Так, 7 августа с больным было 4 приступа; 8 августа—2; 10 августа—1; 11 августа—1; 20 августа—2; 22 августа—1;
13 В. М. Бехтерев
225
23 августа — 1. Большею частью припадки выражаются сердцебиением, страхом и сопровождаются помрачением сознания.
С конца aerycia больной начал вместе с бромидами принимать Inf. digitalis в следующей микстуре: Inf. herbae digitalis е. gr. vjjj—vjjj—Svj Natrii bromati 3]'jj, Aq. laurocerasi 3jj , no 4 ложки в день.
Приступы вначале совершенно приостановились: лишь 7и8 сентября случилось по одному приступу сердцебиения без потери сознания. С9 сентября ввиду появления этих слабых припадков сердцебиения lnf, digitalis в бромистой микстуре заменен Inf. Adonis vernalis в следующей пропорции: Inf. Adonis ve^naii; зЭ — svj, а с 14 сентября в пропорции эй —gvj. С тех пор приступы прекратились совершенно. Больной уехал от меня из Казани пред окончанием навигации в сентябре без припадков. По возвращении домой в Нижний Новгород больной снова стал подвергаться припадкам, но лишь очень слабым, выражавшимся мимолетным чувством страха, большею частью даже незаметным дл-я окружающих. Впрочем, изредка с больным случались и более сильные приступы. Хотя больной и дома продолжал принимать бромиды с Inf. Adonis vernalis, но, повидимому, он не мог пользоваться хорошим препаратом последнего. Это выяснилось весной 1891 г., когда больной проводил поочередно по 4 недели в Казани к по 4 недели в Нижнем Новгороде.
В Казани его приступы всегда прекращались, при переезде же в Нижний Новгород они снова появлялись. Ввиду этого 17 июня, когда больной вновь приехал ко мне в Казань, ему предложено остаться в Казани на более долгое время. Незадолго до этого с больным было несколько сильных приступов с падением, а именно: 12 и 26 мая и 4 июля. Кроме того, за тот же период времени было 3 припадка, выражавшихся страхом и сердцебиением. Больному назначено попрежнему Inf. Adonis vernalis из эjj — Jvj с бромистыми препаратами и кодеином по 6 приемов в день. 19 июня появился лишь слабый приступ, состоявший из сердцебиения и страха без потери сознания. Приемы микстуры увеличены до 8 раз в день. С этих пор припадки прекратились, и больной вернулся домой здоровым. Здесь опять возобновились припадки, выражавшиеся лишь страхом и сердцебиением. С новым назначением микстуры из Inf. Adonis vernalis э j j — Svj, Kalii bromati g j J j и t-rae Convallariae majalis g j j j no 6 — 8 столовых ложек в день припадки вновь прекратились.
Приведенные наблюдения представляют собою убедительные примеры успешного лечения эпилептических приступов с помощью Inf. Adonis vernalis, в соединении с бромистыми препаратами. Я не привожу здесь других наблюдений подобного же рода, так как полагаю, что и приведенных достаточно для того, чтобы составить понятие о действии смеси на приступы падучей. Замечу здесь, что в некоторых случаях я видел успех и от назначения Inf. digitalis в смеси с бромистыми препаратами; но ввиду того, что Digitalis обладает кумулятивным действием, затрудняющим продолжительное его применение, я большею частью предпочитал в своих случаях назначать Inf. Adonis vernalis, тем более что успех при лечении Inf. digitalis, по моим наблюдениям, представляется менее верным, чем при лечении Adonis vernalis. Присоединение к вышеуказанной смеси кодеина я нахожу также крайне полезным при лечении падучей и думаю, что и другие опиаты совместно с бромом могут быть применяемы при этой болезни с большим успехом.
О ХОРЕИЧЕСКОЙ ПАДУЧЕЙ
Отношение между пляской и общими судорожными неврозами, как истерия и падучая, до настоящего времени представляется еще весьма мало изученным. Правда, уже давно известно, что существует известная связь между истерией и обыкновенной пляской, которую в последнее время стали называть пляской Сиденгама, в отличие от хронической пляски Гентингтона. Так, нередко у больных, страдающих пляской Сиденгама, а равно и у их родителей, удается обнаружить те или другие проявления истерии. Как известно, многие авторы различают даже особую форму под названием истерической пляски.
Далее, в тех случаях пляски, где не удается обнаружить явных проявлений истерии, иногда можно найти гемианестезию и яичниковую болезненность даже у детей \ а по учению Шарко и его школы, эти явления, как известно, должны быть отнесены к постоянным признакам истерического' невроза. Наконец, и у родителей лиц, страдающих истерией, можно иногда отметить существование пляски. Словом, имеющиеся в настоящее время сведения по этому вопросу говорят, безусловно, в пользу прямого и наследственного отношения многих случаев пляски Сиденгама к истерии.
Напротив того, по отношению к другой форме пляски, известной под названием хронической пляски Гентингтона, наши сведения в указанном отношении еще весьма скудны. Повидимому, однако, здесь уже нельзя установить строгого соотношения между пляской и истерией. По крайней мере исследование доктора Гюэ 2 привели в этом отношении к отрицательным результатам. Автор не мог обнаружить как в своих, так и в опубликованных до сего времени случаях пляски Гентингтона, никаких указаний на истерическую наследственность. Точно так же и у самих больных не было найдено признаков истерического невроза. Расстройства чувствительности при хронической пляске наблюдаются лишь исключительным образом. Они были указаны лишь в небольшом числе наблюдений и к тому же в этих случаях дело идет о таком ослаблении умственных способностей у больных, что
1 MarieP., Progr. med., 1896, p. 39.
2 E. Hiiet, De la choree chronique, Paris, 1883 — 1889.
}:>*
ГЛ
представляется довольно трудным решить с точностью, в какой мере чувствительность была поражена.
Что касается до отношения пляски к падучей, то этот вопрос представляется еще более темным; в особенности наши сведения недостаточны об отношении хронической пляски к падучей.
Однако исследования доктора Гюэ доказывают наследственное отношение падучей и хронической пляски, так как в семьях больных, страдающих хронической пляской, автор неоднократно находил существование падучей. Так, в одном из наблюдений мать больного, страдавшая пляской, была в то же время в течение многих лет падучной. В другом наблюдении пляска существовала в материнской линии, а падучая в отцовской. Что касается самих больных, то иногда у них были наблюдаемы приступы падучей. Однако случаи подобного рода отличаются редкостью.
Из 16 больных доктора Гюэ в двух случаях приступы падучей были за несколько месяцев или несколько лет до начала болезни. У одной больной подобные приступы, повидимому, существовали почти в течение 18 месяцев перед началом ее болезни. Они прекратились вместе с поступлением больной в Сальпетриер и во время всего ее пребывания в больнице, в течение почти 10 месяцев, они не обнаруживались. У другого больного падучные приступы предшествовали за два года появлению пляски и продолжались в течение многих*лет. Они исчезли около трех лет назад. Следует заметить, что у(обоих больных была поздняя падучая. У больного Гофмана также существовало совпадение падучей с хронической пляской; но в этом случае падучая появилась рано, еще в детстве, вскоре после потрясения, пляска же обнаружилась несколько лет спустя, но много ранее, чем обыкновенно появляется хроническая пляска, так как болезнь начала обнаруживаться еще в возрасте, когда больной посещал пшколу.
Один больной Гофмана х заслуживает особого внимания в отношении занимающего нас вопроса. Он сделался падучным спустя много лет после начала своей пляски, тогда как ни его мать, ни обе его сестры никогда не страдали пляской, но получили позднюю падучую. Должно, однако, заметить, что некоторые особенности этого случая выделяют его из ряда других наблюдений с хронической пляской вследствие существования атрофии некоторых мест на руках, благодаря чему многие авторы не считают этот случай за случай действительной пляски.
Что касается до более тесного соотношения между пляской и падучей во всех вышеуказанных случаях, где обе болезни существовали одновременно, то мы не замечаем в этом отношении ничего особенного. Соотношение обеих болезней друг с другом ничем существенным не проявляется и можно сказать, что и та, и другая болезнь существует в одном организме как бы независимо друг от друга.
1 Hoffmann, Virch. Arch., 1888, Bd. Ill, S. 832.
228
Но в литературе указывалось и на возможность совместного существования пляски и падучей, где напряженность одной болезни обратно пропорциональна напряженности другой болезни. Случаи такого рода на мой взгляд представляют исключительный интерес. К такому сочетанию пляски в виде судорожных подёргиваний в разных частях тела с падучей должен быть отнесен, между прочим, нижеследующий случай
Больной М., 21 года, крестьянин, православного вероисповедания, х олост, родился в Новгородском уезде (Новгородской губернии), где живет все время и занимается печением баранок (бубликов).
Отец больного более 20 лет злоупотребляет спиртными напитками, бывая почти ежедневно пьяным; он вспыльчив и раздражителей; ему теперь 46 лет и он всегда был здоров. Мать больного страдает катарром желудка и имеет 43 года от роду. У больного в живых одна сестра, 17 лет, которая никогда ничем не страдала. Кроме того, у него была еще одна сестра и один брат, которые умерли в раннем детстве.
В детстве больной не помнит никаких болезней, если не считать воспаления подчелюстных желез, наблюдавшегося у больного на 4-м году жизни в течение 2 — 3 недель. Судорог у больного не было. На котором году стал ходить и говорить, ие помнит, но знает, что не позже других детей. На 7-м году больной поступил в сельское училище, в котором хорошо учился и которое окончил с наградой. Будучи в училище, больной в наказание был поставлен на колени и в это время у него обнаружились подергивания в руках, которые потом распространились на все туловище. Было это в 1888 г. Год спустя (в 1889 г.) после непродолжительного сна на солнце у больного наблюдался эпилептический приступ. По словам матери больного, у него бывали эпилептические приступы по ночам еще с детства, но они бывали так редко, что на них не обращали внимания, да и сам больной о них ничего не знает. После приступа в 1889 г. следующий был через год, затем приступы стали повторяться чаще: раз в 3 месяца, позже раз в месяц, а с 1894 г. они бывают почти каждую неделю, притом чаще во сне, но больной о них знает, так как чувствует себя на следующее утро разбитым. Перед приступом начинаются подергивания, которые быстро усиливаются и приступ обыкновенно наступает во время сильных подергиваний. В виде ауры больной при сильных подергиваниях начинает чувствовать особое ощущение в пальцах левой руки и ноги, причем, по словам больного, ему иногда удается предупредить и даже остановить развитие приступа или перетягиванием конечности, или растиранием ее. Если же эти средства не помогают, то ощущения, начавшиеся в пальцах, распространяются выше и доходят до головы, после чего начинается уже самый приступ судорогою в той конечности, где впервые начались ощущения, а затем больной уже теряет сознание. После приступа хореические подергивания на некоторое время ослабевают.
Что касается подергиваний, то они с течением времени стали все более и более усиливаться и заняли постепенно все большую и большую часть тела; так, сначала они были в руках, затем появились в ногах, лице и, наконец, захватили все тело. Особенно тяжелы для больного подергивания в ногах, так как они нередко служат причиною падения больного: «Точно кто-то ударяем меня под колена, и я должен падать», говорит больной. Кроме того ; у больного наблюдаются частые головокружения с кратковременными потерями сознания. С жалобами на подергивания во всем теле, на припадки и головокружения с потерею сознания больной и поступил в клинику 24 января 1896 г.
Что касается образа жизни больного, то он всегда был умеренный. Больной онанизмом не занимался (исключая короткого времени в школе), abusus in Baccho et Venere (излишки в увлечении вином и женщинами) не было, переутомления также не было, наконец, больной отрицает и сифилис. Условия жизни также были удовлетворительны.
229
Status p г a e s'e π s. Больной среднего роста, умеренного тело сложения и питания. Кожа чиста, нигде нет ни язв, ни сыпи, ни рубцов, ни пятен. Костная и мышечная система развиты хорошо и правильно. Суставы функционируют правильно, железы не прощупываются. Череп асимметрии не представляет: бугров, рубцов и чувствительных мест на нем нет. Позвоночник по форме и подвижности отклонений от нормы не представляет; позвонки при постукивании молотком нечувствительны. Паренхиматозные органы (печень, почки, селезенка, желудок и кишечник) при исследовании видимых отклонений от нормы не обнаруживают. Сердце работает хорошо, тоны чисты, пульс правильного ритма, хорошего наполнения, 76 в минуту. Легкие ни при перкуссии, ни при аускультации не обнаруживают ничего ненормального. Психическая сфера представляется нормальной. Что касается речи больного, то нужно отметить, что она по поеменам прерывается и больной как бы заикается. Прерывание речи по времени совпадает с подергиваниями в других частях тела.
Походка больного вне судорог вполне правильная, но во время судорог и подергиваний в ногах больной часто падает, как будто кто-то ударяет его под колени. Пассивные движения совершаются правильно. Тонус мышц несколько повышен. Сила мышц правой руки 90 ф., левой —■ 80 ф. Симптом Ромберга отсутствует, в пальцах вытянутой руки замечается слабое дрожание. У больного существуют подергивания во всем теле, которые появляются беспрерывно, непроизвольно и порывисто; подергивания эти усиливаются при волевых движениях и при волнении больного и характеризуются неправильным и быстрым характером; при психическом возбуждении они до того усиливаются, что больной лишается возможности ходить, сидеть и стоять. Кроме этих подергиваний, у больного наблюдаются эпилептические приступы, сопровождающиеся общими судорогами, длящиеся от 5 до 7 минут, в виде ауры наблюдается усиленное сердцебиение. Кожные рефлексы повышены, причем они особенно резко выражены, если больной не ждет прикосновения к телу. Сухожильные рефлексы также повышены резко. Особенно резко повышены коленные рефлексы. Клонус стопы отсутствует.
Чувствительность больного во всех ее видах не изменена: она одинакова на симметричных местах, нигде нет ненормальных ощущений, равно как нет нигде болей. Мышечное чувство на нарушено.
Со стороны вазомоторной системы нужно отметить существующий у больного дермографизм.
После применения гипнотических внушений больной спит спокойнее и подергивания в течение многих часов довольно значительно ослабевают. Нужно еще добавить, что во время долговременного пребывания больного в клинике в общем его состояние представлялось стационарным.
Мы видим, таким образом, что наше наблюдение не похоже на большинство других опубликованных до сих пор случаев совпадения пляски с падучей. Здесь дело идет не о простом совпадении одной формы с другой, а о действительном соотношении между падучей и пляской в виде судорожных подергиваний членов, выражающемся, между прочим, и в течении болезни, так как подергивания постепенно нарастают в своей степени впредь до развития приступа падучей, после же последнего они на некоторое время почти совершенно прекращаются, а затем начинают снова нарастать до следующего приступа падучей. Если приступ падучей вследствие той или другой причины долго не наступает, то и подергивания членов усиливаются до необычайной степени. Самый приступ является как бы простым усилением этих подергиваний до настоящих судорог, сопровождающихся потерей
<5ЧП
сознания. Более сильные приступы падучей обыкновенно на более долгое время и более резким образом облегчают и судорожные подергивания у больного.
Таким образом, представляется несомненным, что пляска в виде судорожных подергиваний здесь не только осложняет падучую, но она имеет и причинную связь с ней, являясь как бы выражением одного и того же раздражения в центрах.
Ближайшее рассмотрение случая нас убеждает в том, что основной формой здесь является падучая, которая существовала у больного еще с раннего возраста и некоторое время не сопровождалась пляской, затем падучая на время затихла до тех пор, пока больной не получил, лежа на солнце, нового приступа. Незадолго до последнего после сильного нравственного волнения у больного впервые появились судорожные подергивания и толчки в теле, которые со временем стали постепенно усиливаться.
В силу тесного соотношения судорожных подергиваний с приступами падучей мы и называем болезненное состояние нашего больного хореической или плясковой падучей (epilepsia chronica). Наш больной, таким образом, представляет как бы особый вид падучей, причем соотношение пляски в виде судорожных подергиваний с падучными приступами в данном случае может быть вполне уподоблено тому отношению, которое представляют постоянные судороги к приступу падучей при постоянной частичной падучей, описанной Кожевниковым.
Возникает теперь вопрос, как мы можем объяснить себе сочетание судорожных подергиваний с падучей, иначе говоря, имеем ли мы здесь действительно заболевание с одной патологоанатоми-ческой основой или же мы имеем в нашем случае какое-либо иное соотношение двух болезненных форм.
В этом отношении нужно прежде всего иметь в виду, что па-дучные приступы, как теперь выяснено путем обстоятельных и точных исследований, произведенных в заведываемои мною лаборатории докторами Тодорскимг и Боришпольским2, сопровождаются активным приливом крови к мозгу, обусловленным, повидимому, сильным сжатием сосудов тела. Таким образом, приходится допустить, что в основе падучных приступов лежит активная гиперемия мозга, вызванная вторичным образом, вследствие общего сосудодвигательного спазма в теле. Столь резкие изменения мозгового кровообращения, лежащие в основе падучных приступов, без сомнения, не остаются вполне бесследными для питания ткани мозга, в силу чего в известных случаях могут происходить более стойкие изменения ткани мозга, особенно мозговой коры и мозговых оболочек, которые и могут служить при-
1 Тодорский, Врач, 1895, № 25. Медицинское обозрение, 1894, № 10; см. также Бехтерев, Неврологический вестник, 1894, т. II, в. 3.
2 Боришпольский, Обозрение психиатрии, 1897, № 6; его же диссертация, СПБ, 1896.
2.11
чиной более или менее постоянных судорожных подергиваний в различных частях тела, напоминающих хореические судороги. Вместе с этим не трудно объяснить и колебания в степени этих судорог, в зависимости от развития падучных приступов. Естественно, что вместе с постепенным нарастанием возбудимости мозговой коры, предшествующим развитию падучного приступа, хореические судорожные подергивания постепенно усиливаются как в своей интенсивности, так и в отношении распространенности. С наступлением же падучного приступа, когда возбудимость мозговой коры, как показывают физиологические исследования, более или менее значительно падает, и хореические подергивания временно прекращаются совершенно или же более или менее значительно ослабевают.
О МАЛОИЗВЕСТНЫХ РАННИХ СИМПТОМАХ СПИННОЙ СУХОТКИ
Известно, в какой степени важно в практическом отношении распознавать спинную сухотку в ранних ее стадиях. Ввиду этого уже давно стали обращать внимание на ранние проявления спинной сухотки, которая чаще всего характеризуется классической триадой симптомов —отсутствием сухожильных рефлексов, молниеносными болями и вялостью реакции и сужением зрачков на свет. Известно, впрочем, что это сочетание ранних симптомов далеко не отличается постоянством и, с другой стороны, в числе ранних проявлений спинной сухотки отмечается также и много других симптомов, встречающихся,'однако, с меньшим постоянством, нежели только что указанные.
Ввиду этого вышеуказанное классическое сочетание симптомов в практическом отношении представляется наиболее важным. Надо, однако, заметить, что из числа трех поименованных симптомов боли представляют собою субъективное явление и притом не отличающееся большим постоянством в своей силе, что затрудняет иногда должную оценку этого симптома. С другой стороны, и вялость реакции и сужение зрачков нередко бывают выражены в такой степени слабо, что представляются трудно уловимыми. Ввиду этих данных изучение других ранних признаков tabes представляется, вообще говоря, в высшей степени важным. В этом отношении уже Шар ко и другими авторами были указаны некоторые из ранних признаков tabes (например, кризы, артропатия и пр.); но эти признаки отличаются уже много меньшим постоянством, нежели предыдущие.
В этом отношении уже много лет тому назад я обратил внимание на своеобразное соотношение при tabes некоторых кожных рефлексов, особенно брюшного и надчревного, и сухожильных коленного и ахиллова рефлексов. Первые обыкновенно долго сохраняются при tabes и нередко даже в начальных стадиях болезни представляются повышенными, тогда как сухожильные рефлексы нижних конечностей (коленный и ахилловый), как известно, уже в начале болезни совершенно потухают г.
1 На усиление брюшных рефлексов указал, между прочим, и О. Ро-зенбах в 1892 г., приписывая ему известное диагностическое значение (см. Zbl. f. Nervenheilk., 1894, № 4).
233
Вышеуказанное соотношение было в последнее время предметом обстоятельного исследования со стороны ассистента нервной клиники доктора II. А. Останкова1, который на основании целого ряда собранных им наблюдений в заведываемой мною клинике пришел к выводу, что рассматриваемый симптом встречается нередко в весьма ранних периодах tabes и отличается довольно значительным постоянством. Ввиду этого при диагнозе ранних степеней tabes, между прочим, необходимо обращать внимание не на одни только сухожильные рефлексы нижних конечностей, но и на совместное состояние кожных, особенно брюшных и надчревных рефлексов. Значение этого симптома особенно ценно ввиду того, что мы имеем дело с объективным явлением и притом легко уловимым, с другой стороны, ввиду того, что уже одно сочетание усиления кожных и угасания сухожильных рефлексов нижней половины тела само по себе представляется характеристичным для ранних периодов tabes. Повидимому, такое соотношение вышеуказанных рефлексов может получить также известное значение и для отличия истинной tabes от pseudotabes, но в этом отношении нужны еще особые наблюдения.
Не очень давно доктор Бернацкий2 описал своеобразный симптом при tabes dorsalis, который состоит в том, что поколачивание молоточком по п. ulnaris позади локтевого сгиба представляется во многих случаях tabes безболезненным. Симптом этот, как известно, был наблюдаем и другими авторами при tabes. Некоторыми авторами (Крамер 3, Бедекер и Фалькенберг 4, Гебель 5), кроме того, отмечалось присутствие этого симптома в прогрессивном параличе помешанных и даже в других формах душевных болезней, а Гесс 6 находил его у эпилептиков в послеприпадоч-ном состоянии.
Впрочем, некоторыми из авторов (например, доктором Орханским 7) умалялось диагностическое значение вышеуказанного симптома для tabes dorsalis. В последнее время, однако, наблюдения доктора А. Сарбо8 вновь отмечают диагностическое значение как аналгезии п. ulnaris, так и п. peronei для tabes. Последний нерв автор исследовал в том предположении, что при tabes аналгезия п. peronei, казалось бы, должна встречаться чаще, чем аналгезия п. ulnaris, что, однако, не оправдалось в случаях автора.
1 См. доклад в Научных собраниях врачей С.-Петербургской клиники душевных и нервных болезней, Врач, 1897, № 33.
2 Бернацкий, Analgesie des Ulnarisstammes als Tabessymptom, Neurol. Zbl., 1894, № 7.
3 Cramer, Neurol. Zbl., 1894, № 13.
4 Boedecker und. Falkenberg, Alg. Zschr. f. Psychiatrie, 1896. S. 196.
5 Gdbel, Neurol., Zbl., 1895, № 6.
6 Hess, Neurol Zbl., 1896, № 1.
7 Orchansky, Inaug. Diss., 1895; Neurcl. Zbl., 1895, № 8.
8 S a r b о Л., Neurol. Zbl., 1896, № 8.
23-*
Что касается моих наблюдений относительно симптома Бер-нацкого' при tabes, то я должен сказать, что его обыкновенно удается найти в большинстве затянувшихся случаев, но в ранних периодах tabes он нередко отсутствует. Как известно, исследовать этот симптом удается не только путем поколачивания молоточком по нерву, но и просто путем придавливания его пальцами к кости. Последний способ, на мой взгляд, во многих отношениях представляется даже удобнее первого.
Исследуя чувствительность нервных стволов к давлению при различных нервных заболеваниях, я, между прочим, уже давно обратил внимание на особенную доступность к сдавливанию у здоровых лиц, кроме п. ulnarisH η. peroneus, также и п. popli-teus в подколенной области. Эта доступность к сдавливанию п. popliteus объясняется анатомическим положением этого нерва в подколенной области, благодаря чему при полусогнутом положении коленного сустава простым надавливанием одного или нескольких пальцев п. popliteus легко придавливается к подлежащей кости, вследствие чего ощущается ясная боль на месте давления, иногда сопровождающаяся эксцентрическими ощущениями, распространяющимися по направлению нерва к периферии. То же явление можно получить и поколачивавшем с помощью молоточка по подколенной области при полусогнутом положении конечности.
Наблюдения, произведенные мною относительно" этого симптома, убеждают меня в том, что при tabes он встречается, повиди-мому, еще с большей частотой, нежели симптом Бернацкого, причем в некоторых случаях я мог наблюдать его появление в ранних периодах tabes, когда у больных еще не обнаруживался симптом Бернацкого. На мой взгляд в этом отношении желательны сравнительные наблюдения на большом числе случаев, что. может быть сделано лишь с течением времени.
Не лишена также известного диагностического значения у табетиков и безболезненность к давлению мышц, особенно нижних конечностей и, в частности, икроножных. Известно, что если те или другие мышцы, например, двуглавую, икроножные и пр., мы сдавливаем между пальцами, то тем самым мы вызываем в них ясное болевое ощущение, которое становится особенно резким, если мы сдавливаем мышцы в расслабленном их состоянии. У табетиков я неоднократно наблюдал резко пониженную болезненность или даже совершенную безболезненность к давлению мышц нижних и даже верхних конечностей, особенно же икроножных. Явление это, наряду с безболезненностью п. poplitei, наблюдается иногда даже в ранних периодах tabes, когда еще не обнаруживается в ясной степени ни нарушения мышечного чувства, ни даже резких изменений кожной чувствительности.
АПОПЛЕКТИЧЕСКАЯГЕМИТОНИЯ
В клинической патологии издавна известен целый ряд судорожных расстройств движения, следующих за развитием геми-плегии; из них наиболее известны контрактуры, гемихорея, атетоз и дрожание. Обычно эти расстройства следуют за развитием гемиплегии лишь спустя более или менее продолжительное время. Лишь изредка некоторые из этих расстройств, как, например, гемихорея, предшествуют развитию инсульта и гемиплегии. Но, хотя в этих случаях гемихорея и предшествует гемиплегии, тем не менее последняя и здесь, как и в предыдущих случаях, составляет основное страдание, которое лишь осложняется судорожными расстройствами. Но есть случаи, в которых гемипле-гия или вовсе не развивается, или имеются лишь слабые явления пареза, тогда как судорожные движения выступают с необычайною силою или уже тотчас же вслед за инсультом, или же спустя известное время после его наступления, когда паретические явления уже в значительной мере ослабели. Во всяком случае судорожные явления в этих случаях составляют главный фон болезненного расстройства в течение всего позднейшего времени. При этом в отдельных случаях даже и явления апоплексии нередко почти совершенно отсутствуют или же представляются лишь слабо выраженными. Эти состояния по их клиническим особенностям мы имеем полное основание выделять от других более обычных форм послегемиплегических судорожных расстройств движения. Ввиду особого интереса этих состояний я приведу здесь несколько наблюдений подобного рода, которые, на мой взгляд, заслуживают внимания. История первого больного, которого я наблюдал еще около восьми лет тому (назад, заключается в следующем.
1. Больной С, 21 года, с наследственным предрасположением; обратился ко мне за советом в сентябре 1891 г. Его отец человек нервный, давно и часто страдает кашлем. Дед больного по матери страдал припадками эпилепсии. Мать сообщила, что с детства С. был крепким, здоровым ребенком. На шестом году С. перенес тиф. Как только он стал от него оправляться, не более 10 дней спустя после того, как встал с постели, он, сидя однажды на стуле, потянулся с него за монеткой, упавшей на пол, но при этом получил тотчас же головокружение и свалился со стула левым боком. Испуганный и сильно кричащий ребенок был осторожно поднят и
236
перенесен на кровать. Несколько минут, по словам матери, он был без па-мя,ти. Тотчас после этого падения окружающие заметили, что у ребенка перекосило рот и опустилось левое верхнее веко, явления пареза в конечностях левой стороны, что, впрочем, опровергается матерью. Первые дни после падения он был настолько слаб, что с трудом мог сидеть в кровати, но потом, приблизительно через 5 — 7 дней, почувствовал себя более крепким и начал ходить, причем никаких объективных изменений в его движениях родители не заметили. По поводу его состояния тогда же родители обращались к врачу, который посоветовал делать теплые ванны; последние делались ребенку в продолжепие^.целого года, причем парети-ческие явления вскоре исчезли
совершенно. Спустя месяц после „^,,____ ___ .....,-._,.^_„,..,
падения у него начало сводить • - ....—».. .
мизинец левой ноги, а потом постепенно это сведение распространилось на всю ногу и на руку, а затем на всю половину тела, причем особенно резкой степени судороги достигли месяцев через 8—12 после происшествия. Через некоторое время судороги вновь ослабели; но затем в 1882 г. тонические мышечные сокращения в левой половине тела сделались крайне интенсивными и на этот раз уже постоянными, благодаря чему больной не мог иначе ходить, как только при помощи костыля. В этом году с целью улучшения его походки сделана была проф. Студенским тенотомия ахиллова сухожилия на левой ноге; улучшения, однако, не последовало. Судорожные сокращения конечностей становились все сильнее и •сильнее и дошли до степени более или менее постоянной контрактуры,что обнаруживается и в настоящее время. Из прошлого больного следует еще упомянуть, что в 1887 г. в продолжение двух месяцев он был на серных водах; но и это лечение не оказало влияния на ход болезни. В последнее время больной перенес инфлюэнцу, после которой судороги еще более усилились и теперь не дают больному покоя и даже доводят его до бессонницы, сопровождаясь ч\г»ством неприятной тяготы в левых членах.
Status praesens. В настоящее время больной (см. рисунок) представляет собою довольно истощенного субъекта 166 см росту с узкой грудью, обхват которой едва достигает 71 см; подкожный жировой слой развит очень слабо: телосложение плохое. Мышечная система развита недостаточно; цвет покровов и слизистых оболочек бледный. Лопатки сильно выстоят, причем левая лопатка и плечо опущены. При первом же взгляде на больного обращает внимание то обстоятельство, что мышцы левой руки и левой ноги, m. sternocleidomastoideus, mm. pectorales, мышцы лопатки и cucullaris слева сильно тонически напряжены и резко увеличены в объеме и выстоят в виде больших выпуклостей, благодаря чему в обыкновенном положении с опущенными руками плечевая часть левой руки более сильно, чем правая, прижимается к грудной клетке, дыхательная экскурсия с левой стороны слабее, чем с правой, m. biceps резко выстоит, предплечье полусогнуто и находится в положении supinatio, кисть согнута в тыльном

Рис. 1.
237
направлении, пальцы сильно пригнуты к ладони за исключением большого, который оттянут в сторону. В таком же тоническом сокращении находятся мышцы бедра, голени и стопы, вследствие чего нога полусогнута в колене, стопа сильно вытянута, тогда как пальцы сильно согнуты. Кроме всего этого, наружный край стопы приподнят и она отведена кнаружи. Левый sternocleidomastoideus хотя и напряжен умеренно, но видимого влияния на положение головы не оказывает. Гипертрофированные мышцы уже при ощупывании отличаются необычайной плотностью,^особенно в сокращенном состоянии. В мышцах правых конечностей никаких вообще аномалий не обнаруживается. В лицевых мышцах изменения в покойном положении заметны слабо, но при разговоре и свисте, в особенности же, если это сопровождается волнением, тоническое сокращение выступает довольно ясно, вследствие чего левый угол рта несколько оттягивается кнаружи и кверху и на щеке получаются морщины, идущие сверху и снаружи вниз и кнутри. При ударах перкуссионным молоточком по лицевым мышцам эти сокращения не усиливаются и болей не обнаруживается; при ударах же молоточком по мышцам левых конечностей получается усиление сокращения и болезненность. При поглаживании рукой по мышцам бедра, голени, предплечья или плеча судорога мышц несколько ослабевает, но никогда не прекращается совершенно.
Вследствие тонического сокращения в мышцах конечностей больной не может вовсе пользоваться своей левой рукой и вместе с тем походка больного сильно затрудняется, так как, благодаря тоническому напряжению мышц, обнаруживается сильная тугоподвижность во всех вообще сочленениях ноги, вследствие чего во время ходьбы больной своеобразно прихрамывает. Само собою разумеется, что больной сильно утомляется при ходьбе и страшно потеет. Надо, однако, заметить, что вышеописанное положение конечностей не отличается постоянством: хотя все вообще упомянутые мышцы левой стороны находятся в беспрерывном тоническом сокращении, но степень этого сокращения бывает, повидимому, не одинакова, причем в одно время преобладающее влияние оказывает одна их группа, в другое время другая, причем преобладают то сгибатели, то разгибатели; это в особенности обнаруживается в мышцах ноги, благодаря чему стопа время от времени и притом довольно быстро, помимо всякого участия со стороны больного, переходит из состояния сильного вытяжения в положение крайнего тыльного сгибания. В последнем случае походка больного еще более затрудняется благодаря тому, что его левая нога становится на пятку. В руке также обнаруживается по временам преобладание других мышц, благодаря чему ручная кисть из состояния тыльного сгибания внезапно переходит в состояние ладонного сгибания. В настоящее время, впрочем, эти самопроизвольные в общем довольно редкие перемены положения ручной кисти стали случаться еще реже, чем в прежнее время. Никаких явлений атетоза в пальцах не обнаруживается. Если насильственно расправить пальцы руки до известного предела, приблизительно до образования прямого угла с ладонью, то, благодаря сильному сокращению ан-тагонически действующих мышц, пальцам можно придавать те или другие положения и они удерживают его продолжительное время без изменения. Если пальцы вывести из упомянутого полусогнутого положения в сторону сгибания, то сгибатели тотчас же берут перевес над разгибателями и пальцы туго складываются в кулак: если же производить тыльное разгибание пальцев, то вслед за выведением их за вышеописанный предел тотчас же берут перевес разгибатели, кисть сильно отклоняется к тылу, причем более резко выступает действие разгибателя II и V пальца. То же явление обнаруживается и при сгибании и разгибании ручной кисти. Обыкновенно то или другое положение конечности без постороннего вмешательства держится от нескольких дней до нескольких недель и даже нескольких месяцев. Причины этой перемены больной указать не может и обыкновенно даже не замечает сё. Никакой периодичности в перемечах положения конечностей не сущестцует. Они происходят быстро, почти внезапно и совершенно неожиданно для больного.
1'.Й
I'lto уверению родителей больного такое тоническое сокращение мышц у него наблюдается только в течение дня, ночью же, когда он спит, такого сокращения нет и движения во всех суставах левых конечностей настолько же свободны, как и в правых. Точно так же и днем состояние напряжения этих мышц неодинаково. В то время, когда больной сосредоточивает внимание на своих больных конечностях, в особенности же при волнении, а также при врачебном осмотре эти сокращения достигают наибольшего напряжения; при отведении же внимания больного судорожные сокращения заметно ослабевают в своей силе. Больной сам рассказывает, что у него бывают такие минуты, когда он забывает о своем страдании и совершенно свободно идет по улице. Но если в это время ему попадется знакомый человек и каким-нибудь ничтожным жестом или взглядом напомнит ему о его страдании, то он далее уже не в состоянии будет продолжать так же свободно свой путь вследствие усиления судороги.
При измерении мышц той и другой стороны получены следующие цифры. Окружность левого бедра на уровне мошонки 41 см, правого—ЗЭ1/» см; на середине бедра слева — 41 см, справа — 39 см, на 3 см выше patellae, левое бедро—29^ см, правое — 28 см. Окружности голеней в верхней части справа и слева по 26 см,на середине голени справа 23 см,слева—211/3см, подмышками справа — 18 см, слева—17 см; кости левой ручной кисти и левой стопы представляются меньше развитыми по сравнению с правыми. При исследовании рефлексов на здоровой правой стороне уклонений от нормы не замечается, на левой же при сокращенных мышцах их получить почти ие удается, но если путем поглаживания или отвлечения внимания больного тоническую судорогу удается несколько расслабить, то коленный и локтевые рефлексы оказываются даже повышенными сравнительно с правой стороной. Временами при ослаблении мышечного сокращения удается даже получить стопный феномен (clonus) слева. При вызывании подошвенного рефлекса слева усиливается клоническое сокращение мышц всей ноги. Другие кожные рефлексы, не исключая и crernaster, получаются слева, как и справа, в одинаковой степени. Идиомышечная сократительность слева заметно повышена. Фибриллярных сокращений нет. Равным образом никаких уклонений в болевой, термической и осязательной чувствительности не замечается. При надавливании на нервные стволы болезненных точек не обнаруживается и мышечные сокращения не усиливаются. Язык не дрожит и не отклонен. Зрачки равномерны, болевая и световая реакция их нормальны. В зрении никаких уклонений нет. При исследовании слуха оказалось, что левым ухом больной слышит очень плохо, впра-вом же слух в пределах нормы. Костная проводимость на левой стороне в области лобного бугра и переднего края теменной кости ослаблена. • При исследовании мышц и нервных стволов гальваническим током обнаруживалось лишь некоторое ослабление гальванической возбудимости мышц; для получения наименьшего сокращения в левых конечностях при замыкании катода всегда требовался более сильный ток, нежели для такого же сокращения на правой стороне. С другой стороны, при исследовании фарадическим током для получения наименьшего сокращения на левой руке почти всегда требовался более сильный ток, нежели для такого же сокращения в мышцах правой половины тела. Так, для получения наименьшего сокращения в мышцах расстояние катушки Дюбуа-Реймона было следующим: на предплечье справа 18,5 см, слева — 18,14 см, на середине бедра справа —18 см, слева —17,3 см; па внутренней части голени справа — 18,3 см, слева — 18 см; при исследовании других органов обнаружен лишь слабый шум при верхушке сердца.
В заключение следует упомянуть, что больной вскоре после нашего исследования заболел катарралыюй пневмонией и легкой степенью дизентерии и должен был слечь в больницу. Оба вышеуказанных процесса сопровождались высокой температурой и бредом, но, несмотря на то, судорожные явления у больного оставались без малейших изменений и по выздоровлении от вышеуказанных болезней остались в том же положении, как обнаруживались и ранее.
239
2. Больной Н., 26 лет, крестьянин, родился в Петербургской губернии, где и жил, помогая своему отцу в хозяйстве все время, впредь до поступления больного в клинику в начале 1897 г. Отец больного здоров, 50 лет, злоупотреблял спиртными напитками и в молодости страдал какою-то венерической болезнью (по словам матери больного, сифилисом). Мать— 50 лет, раньше страдала маточным кровотечением, а теперь страдает припадками головной боли, сопровождающимися чувством жара и холода в теле и повторяющимися каждую неделю. Припадки эти начались у матери 13 лет назад. Кроме того, она слаба, раздражительна и всякое волнение может у нее вызвать появление припадка. У больного 4 брата, все здоровы. Два брата и одна сестра умерли в раннем детстве (от каких болезней, не знает). О здоровье своих родственников больной не имеет сведений.
Раннее детство больного протекало нормально и он считался здоровым ребенком; на котором году стал ходить и говорить, не помнит точно, но ие позже 3-го года. На 7-м году жизни больной осенью (в октябре) отправился босой в соседнюю деревню; возвратившись к вечеру того же дня домой также босым, он заболел какою-то лихорадочною болезнью, сопровождавшейся сильным жаром и потерей сознания в течение 3 суток, причем во все это время и днем и ночью больной кричал, жалуясь на боли и колики во всем теле. На 4-й день жар стал уменьшаться, но в продолжение 4 месяцев больной не мог вставать с кровати. К концу 4-го месяца болезни больной впал в глубокий сон или, вернее, кому, в которой находился целые сутки, так что окружающие считали его умершим. Когда он проснулся, то у него отнялся язык и появилось «параличное состояние в правой руке и правой ноге». Через день речь стала возвращаться. В конечностях у больного был парез, так как, по его словам, он не мог правой рукой поддерживать струг при работе. Через 2 месяца, когда парез уже ослабел, в руке начала обнаруживаться тоническая судорога, благодаря которой рука вытянулась кверху выше головы; спустя некоторое время и ногу стало тянуть и она согнулась в колене. С течением времени нога сделалась более подвижной и выпрямилась, причем больной стал ходить при согнутых пальцах правой ноги; рука же осталась без всякого улучшения. На 10-м году больной стал учиться грамоте и научился писать левой рукой. На 14-м году больной стал заниматься онанизмом, что продолжалось до 20 лет. С женщинами больной до сих пор не имел сношений. Спиртные иапитки начал употреблять последние 2—3 года, но злоупотреблений в этом отношении не было. Лет 5 назад больной заболел какою-то лихорадочною болезнью, сопровождавшейся в течение 2 недель высокою температурой и продержавшей больного около 8 недель в кровати. Года 4 назад он упал с лошади и пролежал около 2 часов без чувств. Года 3 назад у больного была сильная рвота, и с тех пор у него начали являться тошнота и боли в животе после еды, равно как и постоянные запоры.
Но главные жалобы больного заключаются в невозможности владеть правой рукой и ногой, в чувстве сильного «стягивания» этих конечностей, которое усиливается по ночам и во время дурной погоды, и в судорогах в них, особенно при волнении и неприятных известиях, так что больной не может тогда ни стоять, ни сидеть, а только лежать. С этими жалобами больной и поступил 7 февраля 1897 г. в клинику.
Status praesens. Больной роста выше среднего, умеренного телосложения и достаточного питания. Кожа чиста, нигде нет ни язв, ни рубцов,ни сыпи и пятен. Видимые слизистые оболочки розового цвета. Костная система развита хорошо и правильно; суставы функционируют хорошо, железы нигде не прощупываются. Череп представляет небольшую асимметрию, как бы скошен влево; бугров, вдавлений и рубцов на нем нет. В лице заметна небольшая асимметрия, лицо как бы скошено влево, но при волнении нередко у больного развивается род тонической судороги в мышцах правой щеки, вследствие чего она оттягивается кзади и получается резкая носогубная складка с правой стороны.
В обычном состоянии у больного правый угол рта несколько опущен, ио при открытом рте лицо стягивается заметно в правую сторону; мышцы
240
лица сокращаются правильно, ио иа левой стороне эти сокращения лучше выражены сравнительно с правой. Речь больвого немного затруднительна, причём язык, по словам больного, часто «цепляется за зубы» (вследствие судорожных сокращений) и не дает ему говорить.
Походка паретически-спастическая: больной волочит правую ногу, которая не сгибается в колене и как бы приклеена к полу; при походке видно, что больной ступает только на концы пальцев правой ноги, по временам же заметны судороги в правой ноге. При быстрой походке слышно, как больной скребет пол носком правой ноги. Правая йога представляется несколько атрофированной. Размеры правой ноги в окружности бедра 49 см, левой йоги — 49,5 см, размер правой голени 34,5 см, левой— 35,0 см.
Мышцы правой ноги (особенно бедра) напряжены. Нога обычно находится в вытянутом положении вследствие тонического состояния ее мышц, которое по временам заметно усиливается. Судороги замечаются как во флексорах, так и в экстензорах. Благодаря судороге, пассивные движения затруднены в голеностопном и коленном суставах этой ноги, особенно же затруднено сгибание и в коленном суставе. При этих движениях замечается весьма резкая ригидность мышц. Активные движения в коленном, голеностопном и пальцевых суставах правой ноги значительно затруднены. .Мышечная сила правой ноги значительно ослаблена.
Правая рука постоянно приподнята, отведена в сторону и вперед, предплечье пронировано, кисть сжата в кулак, флексирована и отведена, большой палец вытянут, экстензирован и отведен. Мышцы правой руки и правого плечевого пояса напряжены, muse, cucullaris правой стороны также обнаруживает заметное напряжение. Вследствие напряжения мышц правого плечевого пояса правая лопатка отстоит от позвоночника дальше, чем левая: правая отстоит на 11 см, левая на —7 см. Правая рука представляется несколько и притом неравномерно атрофированной (особенно при сравнении с левой). Размер правой руки в окружности плеча 24 см, левой— 26,2 см, размер правого предплечья 25,5 см, левого—26,5 см, но в верхней трети плеча окружность его представляется большей, нежели слева.
Тонус мышц правой руки значительно повышен. Сокращенные мышцы (deltoideus, triceps и пр.) обыкновенно выстоят в виде выпуклин. Пассивные движения совершаются с большим трудом, за исключением плечевого сустава, где движения эти более свободны, особенно вверх. При производ» стве пассивных движений замечается значительная ригидность мышц этой руки. Кисть сжата; чтобы разжать ее, нужно употребить значительное усилие, причем в мышцах предплечья (особенно в экстензорах) появляется судорога. Сам больной пальцы правой руки разгибает с помощью другой руки; особенно трудно согнуть большой палец, который всегда разогнут. Активные движения в правой руке почти невозможны вследствие сильного напряжения мышц. Несмотря на огромное противодействие пассивным движениям в судорожно сокращенных мышцах предплечья и плеча, мышечная сила кисти правой руки значительно ослаблена: динамометр справа показывает 40 кг, на левой руке 105 кг.
Судорога в правых конечностях всегда усиливается при волнении при обращении внимания на больного, при пассивных движениях, при попытках к активным движениям и пр., при этом, как упомянуто, появляется иногда судорожное стягивание и в правой щеке; при других же условиях судорога заметно ослабевает, но вполне, повидимому, никогда не оставляет больного, по крайней мере в бодрственном состоянии; благодаря усилению судорожных сокращений при вышеуказанных условиях, изредка у больного обнаруживаются самостоятельные перемещения судорожно сокращенной правой руки и йоги в пространстве.
Кожные рефлексы сохранены, лопаточный справа резче выражен, нежели слева, ягодичные, яичковые и подошвенный значительно повышены на обеих сторонах.
Сухожильные рефлексы m. bicipitis повышены, рефлексы т. tricipiti s справа вызвать трудно вследствие тонического напряжения, слева ие
16 В. М. Бехтерев
241
|
|
Правая сторона Р.К. |
Левая сторона! Р.К. |
|
|
85 |
85 |
|
|
65 |
75 |
|
|
75 |
75 |
|
|
55 |
70 |
|
|
70' |
84 |
|
|
65 |
65 |
|
|
75 |
75 |
|
|
60 |
60 |
понижен. Коленрый рефлекс повышен на обеих сторонах, но справа сильнее; с ахиллова сухо килия рефлекс также повышен, причем справа получается 3—4 клониче<_ких сокращения икроножных мышц. Попытка вызвать справа клонус стопы сопровождается усилением тонического напряжения в мышцах всей ноги. Механическая возбудимость мышц на правых конечностях значительно повышена. Фарадическая возбудимость дает следующее;
N, medianus ..........
N. radialis ...........
N. ulnaris............
N. peroneus...........
M. biceps............
M. extens, digit, com.......
M. fcex. digit, subl........
M. rectus femoris........
Чувствительность во всех ее видах, не исключая и мышечного чувства, сохранена.
Зрачки расширены, причем правый представляется более широким, чем левый. Реакции световая и аккомодативная сохранены. Функция мочевого пузыря правильная. У больного частые запоры. Со стороны половой функции нужно отметить частые поллюции и эрекции. Со стороны вазомоторной сферы заслуживает внимания резко выраженный дермографизм, причем реакция долго продолжается и по окончании раздражения. Внутренние органы здоровы; атероматоза сосудов нет.
Из субъективных ощущений больного нужно отметить ощущение «стягивания» в правых конечностях, которое усиливается по ночам и во время дурной погоды, равно как и при всяком заболевании больного. Ни головокружений, ни головных болей у больного нет: психическая сфера нормальна.
3. Больная В., 14 лет, уроженка Витебской губернии. 5 лет тому назад, весною, внезапно наступило бессознательное состояние, в то время, когда она шла по полю; бессознательное состояние продолжалось 5 дней. Когда она пришла в себя, то стала замечать стягивание мышц в правой половине лица, в правой руке и правой ноге. Правостороннего же паралича, по ее словам, не было. Степень сокращения мышц в пораженных частях остается за все время одною и тою же, нисколько не ослабевая и не усиливаясь. 3 года тому назад перенесла какую-то лихорадочную болезнь (была лихорадка, головная боль, боль в животе). В детстве никакими болезнями-не страдала.
Сестре больной теперь 7 лет, она совершенно здорова. Оба брата больной (одному было 3 года, другому — 5 лет) умерли от коклюша. Одна сестра больной умерла 2 лет, от какой болезни—неизвестно. Отец больной умер год тому назад от катарра желудка на 40-м году жизни. Мать здорова.
Status praesens. Больная хорошего телосложения. Подкожный жирный слой развит хорошо. Мышечная и костная системы развиты нормально. Правое плечо стоит выше левого. Правая лопатка стоит выше левой и более приближена к позвоночнику. Позвоночник в верхне-груд-
242
ном'своем отделе искривлен вправо. Правую верхнюю конечность держит разогнутою в локтевом и кистевом суставах, а I палец в положении аддукции. Все мышцы ее, особенно плеча, напряжены и выстоят в виде выпуклостей. Желая ослабить тоническое сокращение мышц, она с явным усилием сгибает правую верхнюю конечность левой рукой и прижимает ей в согнутом виде (в локте и кисти) к туловищу.
Правый m. cucullaris и sterno-cleido-mastoideus тонически напряжен. Когда больная стоит, то не чувствует стягивания в правой ноге, хотя видно, что мышцы'ее напряжены более мышц левой нижней конечности. Когда больная стоит на одной левой ноге, то происходит тыльное сгибание правой стопы вследствие тонического сокращения сгибателей голени и поворот ее киаружи. Тоническое сокращение мышц для больной в общем болезненно; во время сна оно будто бы не исчезает. Левый коленный рефлекс выражен резче правого. Рефлекс с ахиллова сухожилия получается только слева. Рефлексы с mm. biceps, triceps, с мышц предплечья справа выражены весьма слабо. Кожные рефлексы надбрюшной и брюшной справа выражены резче; правый подошвенный рефлекс выражен лучше левого. Левой рукой выжимает динамометр 55 кг. Правой рукой сжимать динамометр не может. Мышечная сила в правых конечностях несколько слабее. Вкус, обоняние и слух сохранены. Зрение в порядке. Реакции зрачков на свет и аккомодацию сохранены. Язык отклонен вправо. Чувствительность без изменений.
Во всех трех приведенных наблюдениях имеется много общего между собою как в этиологическом, так и в клиническом отношениях. Прежде всего во всех случаях поражение развилось в детстве еще до 10-летнего возраста и началось во всех трех случаях апоплексией. Эта апоплексия, видимо, была очень слабо выражена в первом случае, так как потеря чувств продолжалась всего лишь несколько минут, значительно более резко выраженной она представлялась во втором случае, где больной пролежал в коме около суток, и еще резче в третьем случае, где больная оставалась в бессознательном состоянии около 5 дней. Во всех трех случаях инсульт развился внезапно, без всяких предшествовавших явлений. Надо, впрочем, заметить, что лишь в последнем случае настоящее расстройство, выражающееся судорожным сокращением мышц, развилось непосредственно вслед за инсультов, в двух же предыдущих случаях вслед за инсультом с самого начала развилось паретическое состояние одной половины тела и затем, спустя некоторое время (в первом случае около одного месяца, во втором случае —около двух месяцев), уже в том периоде, когда паретические явления в значительной мере ослабели и сделались почти незаметными, у больных начинает более или менее быстро развиваться тоническое стягивание пораженных членов, которое постепенно охватывает всю или почти всю половину тела.
Надо, впрочем, заметить, что и в третьем случае паретическое состояние, повидимому, не отсутствует совершенно, так как при исследовании обнаружено, что мышечная сила в пораженных правых конечностях слабее, нежели в левых, причем, например., правой рукой больная не может сжимать динамометр, тогда как левой рукой выжимает его до 55 фунтов.
16*
24:.
Тем не менее интерес вышеприведенных наблюдений заключается в том, что упорная тоническая судорога, распространяющаяся на всю правую половину тела и в особенности резко выраженная в конечностях, обнаруживается при весьма слабых проявлениях параличных расстройств.
В первом случае, например, судорожные явления у больного стали обнаруживаться в том периоде, когда уже заметных явлений пареза не обнаруживалось. Больной, казалось, уже совершенно оправлялся от бывшего с ним инсульта, как вдруг у него начинает судорожно сводить мизинец левой ноги, после чего тоническая судорога распространилась на всю левую нижнюю и верхнюю конечность, а частью и на лицо.
В этиологическом отношении достойно внимания, между прочим, то обстоятельство, что в двух первых случаях инсульт развился после перенесенной болезни общего характера, сопровождавшейся жаром, очевидно инфекционного происхождения. В третьем же случае предшествующие заболеванию условия остались неизвестными.
Что касается собственно тех явлений, которыми обнаруживается болезненное состояние в период наблюдения, то оказывается, что у всех больных дело идет о сильной тонической судороге членов одной стороны, которая распространяется даже и на соответствующую сторону лица. Эта судорога обнаруживается у больных с постоянством в течение бодрственного состояния, ослабевая при отвлечении внимания, при легком поглаживании и усиливаясь при каждом случае обращения с больным, при механическом раздражении мышц (ударами врачебного молотка), при волнениях и т. п. При этом нарастании судороги, естественно, происходит соответствующее смещение членов (отведение и поднимание руки, ротация ее кнаружи и т. п.). Иногда даже вследствие преобладания судорожных сокращений тех или других мышечных групп, развивающегося под влиянием непроизвольных импульсов, происходит самостоятельное перемещение судорожно сокращенных членов и изменение характера контрактуры. Таким образом, стопа переходит из резко ]вытянутого положения в крайнюю степень тыльного сгибания или кисть руки переходит из крайней флексии в положение резкого разгибания и т. п. Эти перемещения и изменения положения членов, однако, случаются лишь по временам в неправильные промежутки времени, более или менее быстро и лишь изредка и вообще имеют мало общего с тем явлением, которое известно под названием атетоза. При этом случае следует упомянуть, что упомянутые смещения обнаруживаются не в пальцах, где чаще всего атетоз, а в других частях конечностей.
В отличие от постоянных пассивных мышечных контрактур, эти тонические судороги отличаются тем, что судорожно сокращенные члены, например, пальцы, кисть руки, стопа и пр., могут быть выведены из своего ненормального положения
244
и некоторое время остаются в новом положении, будучи удерживаемы в нем судорожным сокращением антагонистов, как это обнаруживалось в особенно резкой степени в нашем первом случае. Надо иметь в виду, что в случаях, описанных нами, дело идет сб активных судорожных состояниях, поражающих не одну только группу мышц, а значительное число мышц, нередко действующих антагонистически, и потому то преобладание судороги в той или другой мышечной группе, обусловленное непроизвольными психическими или иными импульсами, приводит иногда к перемещению судорожно сокращенных членов. Эта судорога, повидимому, не вполне исчезает и во время сна. Хотя в первом из наших случаев родители больного и удостоверяют, что во время сна его судороги исчезают, но двое других больных жаловались на чувство мучительного стягивания членов и во сне.
Другое отличие наших случаев от вторичных контрактур — это отсутствие классического положения конечностей.
Как известно, при гемиплегиях вторичная контрактура обычно выражается преобладанием сгибателей верхней конечности, разгибателей нижней конечности. Этим обусловливается всем известное обычное положение конечностей у гемиплегиков, именно: рука приведена и слегка пронирована, предплечье полусогнуто, пальцы сильно согнуты в своих суставах и нередко впиваются в ладонь. В более редких случаях наблюдается преобладание разгибателей и тогда картина представляется иною, рука отведена от туловища и вытянута, плечо обращено кнаружи: и пальцы в пястно-фаланговых суставах разогнуты, а в фалан-говых более или менее согнуты. Рука вообще или пронирована или супинирована. Преобладание же разгибателей над сгибателями Бюшаром отмечалось всего в 5 случаях из 31.
Что касается нижней конечности, то обыкновенно контрактуры выражаются вытянутым положением ноги, причем стопа представляет собою подошвенное сгибание, вследствие чего пере-ст-упание сильно затрудняется. В более редких случаях наблю-да'ют преобладание сгибателей нижней конечности. Что касается мышц лица, то вторичные контрактуры в них вообще составляют крайне редкое явление.
Если мы обратимся к нашим случаям, то мы увидим, что такого постоянного положения членов, как при обыкновенных контрактурах, здесь не бывает. В некоторых случаях даже трудно сказать, преобладает ли у больных тоническое напряжение сгибателей или разгибателей. Так, например, в нашем первом случае нельзя говорить о значительном преобладании судороги одних мышц над другими, так как кисть его руки, находясь обыкновенно в согнутом положении, может быть насильственно выведена из этого положения и тогда остается уже разогнутою и то же самое замечается в отношении стопы. Дальнейшая особенность наших случаев заключается в том, что имеется сильное колебание п силе судороги в зависимости от внимания больных:
245
когда внимание их отвлечено, то и судорога заметно ослабевает, но при малейшем обращении внимания на свои члены, а равно и при волнении судорога тотчас же резко усиливается. Очевидно, что в нашем случае дело идет о настоящей тонической судороге, а не о вторичной стационарной контрактуре тех или других мышечных групп.
Надо заметить, что рядом с сильными тоническими судорогами пораженных членов у больных обнаруживается обыкновенно небольшой парез мышц для произвольных движений. Парез этот легко констатировать, заставив больного, например, сжимать динамометр; при отсутствии же специального исследования на силу произвольного сокращения мышц парез легко может быть просматриваем. Чувствительность ни в одном из наблюдаемых случаев не обнаруживала заметных уклонений от нормы.
Под влиянием постоянного или частого судорожного сокращения некоторые из мышц подвергаются функциональной гипертрофии и выделяются в сокращенном состоянии, как плотные выпуклины, поражающие всякого наблюдателя. Надо, впрочем, заметить, что, наряду с этой функциональной гипертрофией пораженных судорогой мышц, вследствие развития гемипареза в раннем возрасте, в двух наших случаях обнаруживалась атрофия костного скелета конечностей и их мышц, мало или вовсе не пораженных судорогой. Если в третьем случае эта атрофия еще не обнаружилась в ясной степени, то это, быть может, объясняется еще неполным возрастом больной (14 лет) и сравнительно недавним развитием болезни (5 лет). К тому же у этой больной как раз явления пареза оставались все время почти незаметными и вообще значительно слабее выраженными, нежели в двух остальных случаях.
Из других явлений следует отметить повышенную механическую возбудимость мышц пораженной стороны и некоторое повышение фарадической реакции, хотя возбудимость к гальваническому току в первом случае на пораженной стороне оказалась пониженной. Что касается рефлексов, то в случае если удастся тем или иным способом достигнуть расслабления мышц, то обы'чно сухожильные рефлексы представляют некоторое повышение. При напряженном же состоянии мышц их обнаружить не удается.
Так как тоническая судорога одной половины тела в вышет описанных случаях развивалась уже в короткое время вслед за развитием мозгового инсульта и представляла собою наиболее выдающееся болезненное явление, сопровождающее же его; а в некоторых случаях предшествующее ему паретическое состояние членов пораженной стороны представлялось в общем лишь слабо выраженным и вообще мало заметным до той степени, что обычно не вызывало почти никаких жалоб со стороны больных, то я и полагаю правильным называть это состояние апоплектяческой гемктонией.
9,16
Случаи такого рода по их характерны м особенностям, на мой взгляд, следует выделять из общей группы послегемиплеги-чгских расстройств движения, к которым они до сих пор причислялись. Дело в том, что гемиплегии в настоящем смысле слова в этих случаях нет, а вслед за апоплексией обыкновенно обнаруживается лишь относительно слабый парез мышц одной стороны, за которым затем и развивается тоническая судорога членов пораженной стороны, иногда уже в том периоде, когда гемипарез почти исчез или ослабел в значительной степени; в других же случаях, как было в нашем третьем случае, тоническая судорога наступает даже немедленно вслед за апоплексией и совершенн о затемняет присутствие гемипареза, который может быть обнаружен лишь при специальном исследовании больных.
Во всяком случае, уже преобладание судорог над гемипаре-зом вслед за развитием мозгового инсульта указывает на то обстоятельство, что в данном случае дело идет о таком поражении, которое, если и нарушает проводимость пирамидного пучка,' то лишь в слабой мере, главным же образом приводит его волокна в состоянии патологического возбуждения.
Обращаясь к вопросу о патогенезе описываемого страдания, необходимо иметь в виду, что по отношению к гемиплегическим расстройствам движения наибольшей распространенностью пользуется гипотеза Бриссо1, по которому разнообразные непроизвольные двигательные явления у гемиплегиков (контрактуры, гемихорея, атетоз) обусловливаются неполным или полным склерозом пирамидных пучков. Надо, однако, заметить, что для раз вития судорожных явлений, как известно, нет надобности в пере рождении пирамидных пучков, так как достаточно для этого и простого раздражения пирамидных пучков патологическим процессом, как доказывают, например, случаи предгемиплеги-ческих расстройств движения.
Ввиду того, что в наших случаях дело обыкновенно начинается с апоплексии, есть основание думать, что патологический процесс представляется, всего вероятнее, в виде более или менее ограниченного мозгового кровоизлияния, развивающегося, как надо думать, в ближайшем соседстве с волокнами пирамидного пучка, которые, подвергаясь сдавливанию, обнаруживают некоторое нарушение в своей проводимости, главным же образом приводятся в состояние более или менее постоянного патологического возбуждения. Если не всегда тоническая судорога развивается непосредственно вслед за развитием инсульта, а лишь спустя несколько недель, то это, очевидно, объясняется тем, что раздражение проводников в таких случаях начинается лишь в периоде рубцевания мозговой раны, благодаря вовлечению волокон пирамидного пучка в этот процесс последовательного развития мозгового поражения, обусловленного кровоизлиянием Усло-
1 В г is s a u'd, These de Paris, 1880.
247
Еием, способствующим развитию кровоизлияния в двух из наших случаев, могла быть предшествующая лихорадочная, вероятнее всего, инфекционная болезнь, в третьем же случае развитию кровоизлияния могло содействовать продолжительное передвижение и телесное напряжение, вызванное ходьбой в поле.
Само собою разумеется, что при вовлечении в поражение пирамидного пучка должно быть и вторичное перерождение его волокон, которое, вероятнее всего, не остается без влияния на развитие судорожных явлений, но вряд ли это перерождение играет главную роль в патогенезе рассматриваемого страдания.
Что касается до вопроса о локализации кровоизлияния в направлении цереброспинальной оси, то надо иметь в виду прежде всего, что кора мозга не может быть принимаема в расчет, так как раздражение коркового вещества двигательной области, как мы знаем из экспериментальных исследований, вызывает клоническую судорогу членов противоположной стороны. Поэтому является полное основание думать, что в случаях подобных выше описанным, процесс гнездится где-либо по протяжению пирамидного пучка. Локализация в подкорковом веществе мозговых полушарий также представляется мало вероятной ввиду того, что здесь волокна пирамидного пучка образуют довольно обширный венец. Таким образом, принимая во внимание односторонность поражения, распространяющегося на целую половину тела, следует признать, что болезненное гнездо при апоплектической гемитонии должно помещаться на уровне мозговых узлов, по соседству с задним отделом внутренней капсулы, где проходят пирамидные волокна в виде одного сплоченного пучка.
О ПОДЧРЕВНОМ РЕФЛЕКСЕ
До сих пор в числе рефлекторных сокращений мышц живота различали два рефлекса—чревный, или брюшной, и надчревный. По Говерсу \ брюшной рефлекс зависит от сокращения мышц живота, которое может быть вызвано механическим раздражением кожи книзу от края ребер по бокам живота и зависит от спинного мозга на уровне VIII—XII грудного нерва. Надчревный рефлекс, выражающийся втягиванием области epigastrii на соответствующей стороне и выражающийся, по Говерсу, сокращением самих высоких волокон recti abdominis вызывается раздражением боковых частей груди на шестом, пятом, а иногда на четвертом межреберном промежутке и исходит от спинного мозга на уровне IV—VI или VII пары грудных нервов.
Позднейшие авторы обращали внимание главным образом на. чревный рефлекс, несмотря на его меньшее постоянство по сравнению с надчревным рефлексом. Так, профессор Оппенгейм. в своем известном руководстве говорит: «Хотя рефлексы можна вызвать с любого места тела, тем не менее для диагностики особенное значение приобрели рефлексы: 1) подошвенный, 2) чревный и 3) кремастера». Что касается чревного рефлекса, то в том же-руководстве говорится, что «чревный рефлекс исследуют, быстро-проводя по коже живота или подреберья пальцем или тупым инструментом; при этом происходит втягивание живота вследствие сокращения брюшных мышц». Так же приблизительно определяют., этот рефлекс и другие авторы и, между прочим, доктор А. Зла-таров в диссертационной работе, вышедшей в 1900 г. «Для этого. рефлекса характерно сокращение мышц брюшной области, наступающее при поверхностном раздражении этой области»2.
В докладе проф. Штрюмпеля на XXIV собрании юго-западных немецких неврологов и психиатров 3 мы встречаем указание на
1 Г о в е р с, Диагностика болезней спинного мозга.
! Dr. A. Zlataroff, Valeur semiologique de quelques reflexes cutanes et muqueux, 1900; см. также работу: E. G a η a u 1 t, Contribution a l'etude de quelques reflexes dans l'hemiplegie d'origine orga nique, These de Paris, 1898.
8 Prof. A. Striimpell, Ueber das Verhalten der Haut-und Sehnenreflexe bei N erven kran ken, Bd. XXIV. Wandversammlung der Sudwestdeutschen Neurologen und irrenarzte zu Baden-Baden am 3 und. 4 Juni 1899, Neur. Zbl., 1899, Nr. 13, S. 617.
249'
то, что рефлексоггчные области, т. е. области кожи, с которых вызывается рефлекс, при некоторых состояниях расширяются в большей или меньшей степени: «Для обыкновенного сгибатель-ного рефлекса ноги рефлексогенная область у паралитиков распространяется вверх различно, иногда только до области колена, часто до паховой области, но часто и еще много выше. Для брюшного рефлекса рефлексогенная кожная область спускается часто на бедро, для кремастерового рефлекса до области стопы». В нормальном состоянии, повидимому, однако, такого расширения рефлексогенных областей не замечается, если не принимать во внимание, по крайней мере, случаи чрезмерного повышения рефлексов, являющиеся выражением общего невроза в виде истерии или неврастении.
Мои личные наблюдения приводят к тому заключению, что в патологических случаях надчревный рефлекс всегда следует исследовать, наряду с чревным или брюшным, потому, что имеются случаи, где чревный рефлекс отсутствует, в то время как надчревный вызывается легко и может служить объектом исследования для сравнения силы его на той и на другой стороне, например, у гемиплегиков. Но независимо от того, в области живота необходимо иметь в виду еще один рефлекс, на который до сего времени не обращали внимания авторы. Рефлекс этот легко получается при механическом раздражении кожи внутренней поверхности бедра вблизи паховой области (например, быстрым проведением рукояткой молоточка под паховой складкой), результатом чего является сокращение мышц нижнего отдела живота, выражающееся ясным западением надпаховой области живота.
Надо думать, что подчревный рефлекс выполняется при посредстве сокращения нижнего отдела m. obliqni abdominis. Уровень же его передачи должен быть тотчас ниже чревного или абдоминального рефлекса и соответствует нижней грудной области спинного мозга. Ввиду этого, в зависимости от уровня поражения спинного мозга, он может быть сохранен, в то время как чревный рефлекс представляется угасшим, и наоборот. С другой стороны, и в нормальном состоянии этот рефлекс, как я убедился, отличается, повидимому, несколько большим постоянством, нежели чревный рефлекс. А так как отношение его при черепно-мозговых поражениях такое же, как и отношение чревного рефлекса, т. е. и тот, и другой рефлексы в этих случаях представляются большею частью пониженными на стороне гемиплегии, то, очевидно, является необходимым, кроме исследования чревного рефлекса, иметь в виду в каждом случае и исследование описываемого мною подчревного рефлекса.
О НАСИЛЬСТВЕННОМ СМЕХЕ В ВИДЕ ПРИСТУПОВ,
СОПРОВОЖДАЮЩИХСЯ ТОНИЧЕСКОЙ СУДОРОГОЙ
И ЧУВСТВОМ ЩЕКОТАНИЯ В ЛЕВОЙ РУКЕ
В работе под заглавием «Насильственный или неудержимый смех и плач при мозговых поражениях»1 я указал впервые на случаи насильственного смеха при органических гнездовых процессах мозга. Там же я указал на случаи эпилепсии, где приступы стереотипно каждый раз начинались смехом, причем больные впоследствии не сохраняли никакого воспоминания как о приступах, так и о смехе. В этих случаях смех, как я отметил тогда же, служит одним из проявлений эпилептического приступа, являясь автоматическим смехом с характером насильственного.
Недавно мне встретился случай, где при органическом мозговом поражении имелись приступы смеха, вызванного внезапно появлявшимся вслед за тонической судорогой чувством сильного щекотания в левой руке, причем обыкновенно эти приступы не сопровождались даже и помрачением сознания.
Наблюдение это представляет большой научный интерес, а потому я приведу его здесь подробно по истории болезни, любезно составленной для меня доктором П. Останковым.
Элеонора М., 15 лет, уроженка города Одессы. Родилась в срок, доно* шенным ребенком и до года и восьми месяцев отличалась хорошим здоровьем; у ией во-время прорезались зубы, около года и двух недель она начала ходить и в это время говорила уже довольно хорошо. Около года и восьми месяцев однажды без видимой причины Элеонора упала с качелей, вскрикнула, имела рвоту и затем потеряла сознание; ее подняли и перенесли на кровать, причем она сильно в продолжение нескольких часов (от 6 часов утра до 8 часов вечера), кричала, часа через два после падения с качелей в левой руке, а отчасти и в левой ноге появились судороги, которые затихали на некоторое время, после чего возобновлялись с прежней силой. Только через несколько часов к ребенку вернулось сознание, но она не могла некоторое время сказать ни одного слова, хотя и делала видимые к тому усилия. При осмотре врачом было установлено, что вся левая половина тела парализована, больная не видела левым глазом, не слышала левым ухом, повидимому, не могла жевать на этой же стороне, левая рука и нога лежали как пласт. В течение года и двух месяцев не было заметно никакого улучшения,причем ежедневно по нескольку раз в день секундами у больной появлялись приступы тонической судороги в левой руке,причем
1 См. Неврологический вестник за 1893 г. и мои «Нервные болезни в отдельных наблюдениях», в. I, Arch. f. Psych., Bd. XXXVI, 3.
251
левая рука разгибалась во всех сочленениях и отводилась кнаружи. Иногда в левой руке у нее развивалась столь сильная боль, что она, по» словам матери, почти задыхалась от нее. Эти приступы продолжаются с тех пор и до настоящего времени. Во время приступов она не теряет сознания, но, по словам матери, делается какой-то странной, смеется и начинает иногда говорить машинально совершенно не относящиеся к делу вещи. Только, когда было больной 3 года, она постепенно стала видеть, появились движения в руке и ноге, и больная начала опять ходить и говорить; слух также постепенно стал улучшаться. С тех пор улучшение продолжалось, хотя рост парализованной ноги и руки стал несколько замедляться и, кроме того, кисть левой руки осталась с самого начала в сгибательной контрактуре; в такой же контрактуре находятся до сих пор и пальцы той же руки; с тех пор осталось и косоглазие.
На психическом развитии до 10 лет заболевание сказывалось мало. 11 лет больная перенесла корь. 12 лет после усиленных занятий у ней появилась сильная головная боль, сопровождаемая рвотой и повышением температуры, причем зрение в течение трех суток было сильно понижено. Лечивший больную врач, по словам матери, распознал «воспаление мозга», хотя заболевание продолжалось всего несколько дней. Матери кажется, что после этого заболевания психическое развитие пошло медленнее и-вместе с тем медленнее же стало происходить и физическое развитие.
В настоящее время больная жалуется на бывающие с ней по ночам припадки замирания сердца, на судорожные приступы в левой руке, как они выше описаны, причем в последнее время во время судороги в руке бывают ощущения сильного щекотания в ней вместе с явлениями со стороны зрения, так как во время припадков днем у ней темнеет в глазах,а ночью кажется, что комната, где она находится, освещена, как днем; явление это продолжается от нескольких секунд до 1—2 минут. Больная жалуется также, что при усиленных занятиях у ией появляется головная боль. Левым глазом больная видит хуже, чем правым и вовсе не видит снаружи. Кисть и пальцы левой руки совершенно не владеют. В семье отца были нервно-и душевнобольные. Дед со стороны отца умер от порока сердца,, отец тоже страдает пороком сердца, родной брат отца умер душевнобольным, двоюродная сестра отца умерла также душевнобольной. Со стороны матери наследственности нет, лишь брат ма'тери обладал очень плохой памятью и не мог учиться. Алкоголиков не было в родне, не было и падучных.
Наша больная — второй ребенок, всего у матери было трое детей. Старшей дочери было 15 лет, когда она умерла от дифтерита. Мать в детском возрасте была очень хворым ребенком и страдала костоедой, которая перешла к ней будто бы от няньки-кормилицы.
Status praesens. Объем черепа 53 см, кости черепа к постукиванию болезненны, особенно в области лобной кости с левой стороны. Зрачки расширены, световая реакция их выражена резко, левый зрачок несколько шире правого. Если источник света расположен снаружи, т. е. в наружном поле зрения левого глаза, то световой реакции зрачка не получается, но как только освещается внутренняя половина зрачка, реакция световая делается явственной. На правом глазу реакция световая слабее, чем на левом, но разницу при освещении глаза с наружной и внутренней стороны установить труднее. Острота зрения довольно хорошая. Поле зрения при исследовании обнаруживает левостороннюю одноименную ге-мианопсию; обоняние на левой стороне ясно повышено. В лице заметна асимметрия: левый угол рта опущен, левая носогубная складка сглажена, при открытом рте овал его скошен. Язык не отклонен. При ходьбе больной левая нога обнаруживает паретически-спастические явления, причем она волочится по полу носком. Стоять на одной левой ноге больная не может. Левая рука согнута в локтевом сочленении, приведена к туловищу, кисть руки согнута, пальцы полусогнуты, предплечье пронировано. Объем правого плеча 24,5 см, левого — 23,5 см, правого предплечья — 22 см, левого — 20 см; объем правой кисти 19 см., левой— 15 см; объем правого бедра— 33 см; левого—30 см, правой голеии—20 см, левой—19 см. Левая рука и ле-
252
вая нога значительно отстали в развитии от правых конечностей, причем правые конечности длиннее левых на 3—5 см. Особенно недоразвита костная система верхнего плеча елевой стороны, причем левая лопатка значительно меньше правой. Мышечная сила правой руки 45 фунтов, левой—О (по динамометру). Мышечная сила сгибания предплечья, сгибания плеча, приведения и отведения плеча резко ослаблена с левой стороны, супинация же предплечья не получается почти вовсе.
Мышечная сила левой нижней конечности значительно ослаблена. Стопа находится в положении pes equinus.
Если попытаться сделать быстрое разгибание предплечья с левой стороны, то в первый момент встречается препятствие, а затем разгибание совершается легко. При попытке быстро согнуть ногу в левом колене получается тоже в первый момент сопротивление. При исследовании болевой чувствительности обнаруживается довольно значительное повышение ее на всей левой половине тела. Тактильная и температурная чувствительность тоже повышена на всей левой половине.
Механическая мышечная возбудимость не повышена. Сухожильные рефлексы с сухожилия mm. bici pit is и tricipitis, а равно и лопаточно-плечевой рефлекс резко повышены на левой стороне; коленные рефлексы, особенно слева, также повышены, а равно повышен и рефлекс с ахиллова сухожилия. Кожные рефлексы несколько вялы на левой стороне, кроме подошвенного. С левой стороны клонус стопы вследствие ригидности в голеностопном сочленении не получается; симптом Бабинского выражен ясно.
По ночам в последнее время стали происходить приступы страха как бы перед смертью, сопровождаемые сильным сердцебиением.
Что касается вышеописанных приступов с вытяжением руки, то они и до сих пор выражаются тем, что левая рука больной вдруг иа мгновение вытягивается и в ней появляется резкое ощущение щекотания, которое распространяется с руки на область шеи и иногда достигает даже левой половины лица. Под влиянием этого щекотания больная начинает неудержимо и громко смеяться, пока длится это ощущение, что обыкновенно случается с 1/2 минуты и более, после чего она совершенно успокаивается. Помрачения сознания при этом не бывает, последовательного сна также. Припадки такого рода с больной случаются ежедневно в различное время дня без всяких предвестников. Такие припадки случались даже на моих глазах во время приема больной в моем кабинете.
Однажды больная, сидя на кресле, вдруг неожиданно вытянула на мгновение свою левую руку в сторону и начала сильно смеяться и хохотать, хотя рука ее в это время, уже опустилась; на мой вопрос, отчего она смеется, она тотчас же живо ответила, не прекращая смеха, объяснив, что чувствует сильное щекотание в левой руке, при котором она не может воздержаться от смеха, несмотря на все ее желание. Через J/2 или 3/4 минуты смех ее прекратился, она чувствовала себя вновь бодрой и на расспросы отвечала, что во время приступа не испытывала ни головокружения, ни помрачения сознания.
Другой, совершенно подобный же, но лишь более кратковременный приступ случился с больной также на моих глазах во время приема больной. И в этот раз приступ начался вытяже* нием левой руки, ощущением сильного щекотания в ней и неудержимым, как бы судорожным, смехом без всяких явлений головокружения и помрачения сознания. Мать, впрочем, предполагает, что у больной эти приступы иногда сопровождаются и некоторым помрачением сознания, так как будто бы она в это время не всегда отвечает правильно, но сама больная категорически (придает даже слабое помрачение сознания во время своих приступов.
253
Не подлежит,- конечно, сомнению, что у больной дело идет об органическом поражении правой половины мозга в основных его областях, в пользу чего говорит с положительностью левосторонняя гемианопсия с гемиопической реакцией зрачков на свет. Так как вместе с тем у больной имеется левосторонний геми-парез с контрактурой верхней конечности, то является необходимым предположить поражение в области, соседней с задней частью зрительного бугра, где одновременно должен быть поражен как зрительный тракт, так и пирамидный двигательный путь. Вместе с э№м у больной вовлечены в раздражение и чувствительные волокна, что очевидно не только из того, что больная имеет левостороннюю гемигиперестезию, но и из того факта, что во время приступов, наряду с развитием тонической судороги в левой верхней конечности, появляется крайне резкое чувство щекотания, ранее же у больной существовали сильные боли в левой руке, очевидно, являвшиеся симптомом раздражения чувствительных проводников.
Само собою разумеется, что и чувство щекотания в левой руке, ныне наблюдаемое во время приступов и приводящее к судорожному смеху, также обусловливается временным раздражением со стороны мозгового поражения чувствительных проводников, распространяющимся на рефлекторные центры мимических движений, которые, как я показал впервые в других своих работах, расположены в зрительном бугре1.
Самые приступы смеха в данном случае имеют несколько другой характер, нежели в ранее описанных мной и другими авторами случаях мозговых поражений, при которых наблюдался насильственный Смех. Там дело шло о неудержимом смехе, который, правда, развивался насильственным образом, но в развитии его обыкновенно играл роль тот или другой, хотя бы и незначительный, психологический момент, тогда как в вышеприведенном случае развитие смеха происходило рефлекторным путем под влиянием внезапно развивающегося чувства щекотания в левой руке и, очевидно, имело тот же судорожный характер, каким отличается смех, наблюдаемый при обыкновенном щекотании человека.
Хотя приступы в данном случае начинались тонической судорогой левой верхней конечности и, по словам матери, иногда сопровождались даже помрачением сознания, что, впрочем, решительно отрицается самой больной, но так как смех у нее развивался уже по прекращении тонической судороги в левой руке, когда в ней появлялось ощущение сильного щекотания, всегда ясно сознаваемого больной, то этим самым данный случай, очевидно, отличается и от эпилептических приступов с насильственным смехом, который обыкновенно больными не сознается.
1 Бехтерев, Об отправлении зрительных бугров у животных и челопека, Вестник психиатрии, 1885; Virch. Arch., 1887,
254
ОБ АКРОМИАЛЬНОМ ИЛИ ВЕРХУШЕЧНОМ-РЕФЛЕКСЕ1
Исследование рефлексов верхней конечности далеко не полу-, чило такого диагностического значения, как исследование рефлексов нижних конечностей, главным образом благодаря непостоянству их в нормальном состоянии 2. Но,, несмотря на то,, в патологических состояниях при повышении рефлекторной возбудимости рефлексы верхних конечностей выступают часто с необычайным постоянством и представляются более или менее резко выраженными. Даже такие рефлексы, которые обычно не наблюдаются в нормальном состоянии, как известно, обнаруживаются с ясностью в патологическом состоянии, в силу чего они могут получить то или другое диагностическое значение. Руководясь, этим, я хотел бы обратить внимание на один рефлекс, который в подходящих случаях может быть вызван ударами врачебного молоточка по акромиальной части надплечья и по ргос. сога-. coideus. Этот рефлекс обыкновенно состоит из легкого сгибатель-ного движения предплечья, иногда легкого поворота руки внутрь, в отдельных же случаях, при особенном повышении этого рефлекса я наблюдал при этом и сгибательное движение пальцев верхней конечности.
/Рефлекс этот, очевидно, обусловливается сокращением m. coraco-brachialis. и т. bicipititis, короткая головка которого, срастаясь с m. coraco-brachialis, начинается вместе с последним от ргос. coracoideus ключицы, длинная же головка начинается от верхнего края сочленовой поверхности лопатки. Но в известных патологических случаях, сопровождающихся резким повышением рефлекторной возбудимости, рефлекс этот может распространяться и на другие мышечные группы (например, сгибатели пальцев).
Судя по анатомическим данным, рассматриваемый рефлекс должен быть причислен к периостальным рефлексам. Как упомянуто, он вызывается чаще всего при повышении рефлекторной воз-
1 Из сообщения, сделанного в заседании научных собраний врачей клиники душевных и нервных болезней за 1901 г.
2 Литература вопроса о рефлексах верхних конечностей приведена, между прочим, в статье Л. Мора: Zur Physiologie und Pathologie der Sehnen-phanomene an den oberen Extremitaten, Dtsch. Zschr. f. Nervenh., 1901, Bd. XIX, Hft. 2-4.
255
•будимости, особенно при органических гемипарезах и гемиплегиях, а равно и при атрофических параличных состояниях, сопровождающихся повышенною рефлекторной возбудимостью, как, например, при атрофическом боковом склерозе, при функциональных же гемиплегиях повышения этого рефлекса обыкновенно не наблюдается.
При исследовании вышеуказанного рефлекса нужно иметь в виду, что при многократном повторении удары врачебным молоточком по акромиальнои части надплечья иногда сопровождаются ясными болезненными ощущениями. В устранение этого, удары молоточком для вызывания рефлекса можно производить не непосредственно по акромиальнои части надплечья, а через плесси метр.
О ЧУВСТВИТЕЛЬНОСТИ К ПОСТУКИВАНИЮ СКУЛОВОЙ ДУЖКИ КАК ПРИЗНАКЕ ОРГАНИЧЕСКОГО ПОРАЖЕНИЯ ОСНОВНЫХ ЧАСТЕЙ МОЗГА И ЕГО ОБОЛОЧЕК
Как известно, чувствительность черепных покровов головы к постукиванию может давать в некоторых случаях указание на существование в полости черепа и позвоночника органического болезненного процесса. Еще в работе «О звуковой проводимости и постукивании костей черепа и позвоночника» \ относящейся к этому предмету, я обратил внимание на тот факт, что если метод простого постукивания молоточком по костям черепа и позвоноч ника в тех или других случаях мозговых поражений оказывается недостаточным, то нередко местную болезненность при постукивании черепа и позвоночника обнаруживает применяемый мною метод глубокого постукивания, т. е. постукивания, производимого молоточком через плессиметр с большею силою. Применяя тот или другой метод постукивания при исследовании своих больных, я убедился, что в случаях мозговых поражений, лежащих в основных частях мозга и его оболочек, особенной чувствительностью к постукиванию отличается скуловая дужка. Иногда даже тщательное обстукивание головы при глубоких мозговых процессах не дает никаких указаний на местную болезненность черепных покровов и только постукивание скуловой дужки на стороне поражения вызывает легкую чувствительность, а при многократном постукивании даже и болезненность. В других случаях можно открыть болезненность при постукивании скуловой дужки на обеих сторонах, но на стороне поражения она всегда выступает сильнее.
Нередко вместе с тем чувствительность к постукиванию обнаруживается во всей вообще височной области черепа; но и в таких случаях обычно постукивание скуловой дужки представляется резлО болезненным. Как было уже упомянуто выше, ос-бенно часто удается отметить чувствительность или болезненность при постукивании скуловой дужки в случаях поражения основных частей мозга, например, при сифилитических поражениях с характером менинго-энцефалита и при поражениях сосу-
1 В. Бехтерев, Вестник неврологии, 1894,
J7 В. м. Бехтерев
257
дистого происхождения. Иногда даже в старых патологических процессах в основных частях мозга путем постукивания скуловой дужки удается отметить более или менее заметную болезненность; в более же свежих случаях она всегда выступает с ясностью.
Мне кажется, что эта особенная чувствительность скуловой дужки при патологических процессах, локализующихся в основных частях черепной полости, может объясняться тем, что скуловая дужка, упираясь своими концами в две области черепа, при постукивании вовлекает в сотрясение довольно' значительную часть основания черепных покровов.
Во всяком случае описываемый мною признак может иметь известное диагностическое значение, так как, опираясь на него, мне приходилось иногда открывать гнездный болезненный процесс мозга в таких случаях, в которых остальные симптомы могли быть истолкованы в пользу разлитого или даже функционального расстройства. Особенного внимания заслуживает исследование чувствительности к постукиванию скуловой дужки у лиц, у которых подозревается сифилитическое поражение основных частей мозга и его оболочек.
PSEUDOMELIA PARAESTHETICA КАК СИМПТОМ
ЧЕРЕПНО-МОЗГОВОГО ПОРАЖЕНИЯ В ОБЛАСТИ
N. LENTICULARIS
После того как мной был описан в особой статье * симптом, выражающийся чувством ложного положения конечности у больного с поражением боковых частей нижнего отдела продолговатого мозга и переходной области между продолговатым и спинным мозгом, у которого имелись вместе с явлениями двигательного паралича также расстройства общей чувствительности и, между прочим, мышечного чувства, упомянутый симптом послужил предметом исследования приват-доцента П. Останкова (из нашей клиники2) у больного, страдающего сифилитическим менингомиэлитом. В случае, описанном этим автором, дело шло, как и в нашем случае, о поражении шейного отдела спинного мозга с участием и нижнего отдела продолговатого мозга. Этот интересный случай в отношении проявления вышеуказанного симптома во многом предствлял аналогию с первоначально мной описанным случаем, причем интересующий нас симптом распространялся на обе руки и обе ноги. «Как убедится читатель из нижеизложенной истории болезни, —говорит автор рассматриваемой работы, —ощущения нашего больного представляют большую аналогию с ощущениями больного проф. В. М. Бехтерева. И там, и здесь конечность чувствуется в согнутом положении; и там, и здесь в конечности ощущается чувство тяжести; наконец, в обоих случаях коррекция зрения нисколько не устраняет тягостные для больного ощущения. Поразительно также постоянство ложного ощущения: в течение нескольких недель оно держится у больного, не испытывая почти никаких перемен. Одну особенность мы не можем оставить не отмеченной •—это то, что локализация процесса как в случае проф. В.'М. Бехте-
1 В. Бехтерев, Своеобразное распределение паралича чувствительности и движения при поражении боковых частей нижнего отдела продолговатого мозга и переходной области между продолговатым и спинным мозгом, Неврологический вестник, 1894, т. II, в. 1, стр. 1, см. также: В. Бехтерев, Нервные болезни, 1894, в. 1.
2 П. Останков, Чувство ложных конечностей (pseudomelia paraesthetica) как симптом поражения шейной части спинного и нижнего отдела продолговатого мозга, Обозрение психиатрии, 1904, № 9.
259
рева, так а в нашем очень близка: и там, и здесь задеты шейная часть спинного мозга и прилегающая к нему непосредственно область продолговатого мозга».
Ввиду клинического интереса, возбуждаемого вышеуказанным симптомом, мы приведем здесь давно бывший под моим наблюдением случай черепно-мозгового поражения, при котором равным образом наблюдался тот же самый симптом. Интерес этого случая увеличивается тем, что наблюдение это проводилось вплоть до вскрытия.
Больной В., 38 лет, православный, сын псаломщика, занимался письмоводством; вдов. Отец злоупотреблял спиртными напитками, о других родственниках сведений не собрано. Сам больной злоупотреблял спиртными напитками, пил запоем по неделе и больше. С 1873 г. у. него появились головные боли, шум в голове и по временам он стал слышать «голоса». С 1883 г. слуховые галлюцинации стали более стойки, но все-таки выпадали светлые промежутки, когда он бывал свободен от них. В последнее время «голоса» стали слышаться чаще, назойливее, с неприятным характером для больного, так что он даже выработал идеи преследования. В 1884 г. около недели находился в Саратовской психиатрической лечебнице, 8 лет тому назад болел горячкой в продолжение 3 недель; через полгода после этого опять лежал в горячке в продолжение месяца.
В лечебницу поступил 28 января 1887 г. Исследование больного, произведенное на другой день по поступлении в лечебницу врачом последней, обнаружило следующее: В. среднего роста, телосложения посредственного, питания удовлетворительного.
Рефлексы пателлярные и мышечные заметно повышены. Со стороны чувствительности уклонений нет; обоняние, слух, вкус — сохранены.
С психической стороны больной обладал довольно разнообразными и рельефными слуховыми галлюцинациями, которые преимущественно имели неприятный для больного характер, в силу чего настроение духа иногда было раздраженное, больной становился озлобленным, требовательным, придирчивым.
Приблизительно с июля 1887 г. больной начал кашлять, потерял аппетит, питание понемногу расстраивалось, явились ночные поты, в легких обнаружились признаки каверн, хронического бронхита и плеврита.
В феврале J888 г. больной был очень слаб. 29 февраля в истории болезни
отмечено следующее: «Вечером больной внезапно упал с кровати. После
падения у него замечены: парез правого верхнего века, отклонение языка
в левую сторону, расширение левого зрачка, затрудненный выговор слова,
бщий упадок сил с потемнением сознания».
2 февраля с утра «замечено судорожное тоническое сокращение мышц в левой верхней и нижней конечности».
3 февраля «судорожное сокращение в мышцах конечностей левой половины прошло, но наступило параличное состояние этих же конечностей».
6 апреля. Наблюдения, сделанные ассистентом Васильевым в заве-дываемой мною клинике, куда больной поступил в начале апреля 1888 г., состояли в следующем.
Заметная атрофия мышц левых конечностей при резко выраженной общей худобе. Fossa supra-et intraspinalis слева почти не покрыты соответствующими мышцами; заметна также значительная атрофия и вялость в mm. cucularis, pectoralis, во всех мышцах плеча и предплечья (разгибатели атрофированы больше), в mm. interossei; thenar, hypothenar с левой стороны. Левая верхняя конечность в полусогнутом положении, хотя пассивное разгибание возможно, пальцы все полусогнуты. В лице (в области разветвления нижней ветви п. facialis) парез с левой стороны. Ни в языке, ни в губах атрофии не заметно. Движений в левой руке нет и больной только немного приподнимает плечо.
260
Нижняя левая конечность обнаруживает приведение и "сгибание в тазобедренном и коленном суставах, но очень слабые; отведения и разгиба-нья нет. В стопе нет никакого движения. Конечность резко атрофирована, причем m. quadriceps и adductores более сохранены сравнительно с другими мышцами. Мышцы голени спереди все резко атрофированы, промежуток между костями резко углублен. Сзади на голени мышцы еще несколько сохранены, но более атрофированы, чем другие.
Фарадический ток слева соответственно атрофированным мышцам обнаруживает количественное ослабление возбудимости. Постоянный ток на левых конечностях также обнаруживает ослабление возбудимости по сравнению с правыми, но реакция нормального типа. Механическая возбудимость мышц и мышечный валик слева живее, нежели справа; сухожильные рефлексы слева живее; костные рефлексы также слева повышены; коленный рефлекс справа передается на левую ногу, тогда как с левой ноги иа правую подобной передачи не наблюдается. Механическая возбудимость вообще повышена, но больше в аддукторах, в то время, как подошвенный и кремастеровый, а равно и другие кожные рефлексы, в том числе и рефлексы живота, ослаблены на левой половине тела. Мышечное чувство в верхней левой конечности совершенно утрачено; больной совершенно не может определить ее местоположение. В нижней левой конечности мышечное чувство резко ослаблено. Болевая, температурная и тактильная чувствительность сохранены на обеих сторонах.
Измерение конечностей, произведенное в начале апреля 1888 г., дало следующее:
Нижняя часть предплечья правой руки......13,5 см
» » » левой » ......12,5 »
Верхняя часть предплечья правой » ..... 20 »
» » » левой » ......17 »
На уровне m.biceps правой руки.........18 »
» » » » левой руки.........14,5 »
Голень правая .................. 24 »
» левая...................24 »
Бедро правое...................29 »
» левое...................29 »
Верхняя часть бедра с правой стороны......32 »
» » » » левой » ......28 »
Должно отметить у больного существовавшее в периоде развития его паралича особое, чрезвычайно упорное, не покидавшее больного до последних дней жизии ложное ощущение двигающейся руки слева. Больной, не имея возможности двигать своей левой рукой, чувствует, однако, присутствие двигающейся левой верхней конечности, которая иногда закидывается за спину и как бы сдавлена лежащим на ней туловищем, в другой раз лежит на его груди или животе. Больной вполне сознает, что это чувство ложное, так как знает, что его левая рука вполне парализована и тем не менее не может отделаться от своего ощущения даже и в том случае, когда смотрит на свою неподвижно лежащую, парализованную левую верхнюю конечность.
24 июля 1888 г. В. умер. Протокол его вскрытия гласит следующее.
Кости черепа тонки, dyploe плохо развито, местами отсутствует, sulci meningei достаточно развиты. Твердая мозговая оболочка по sinus longitudinalis с наружной стороны ииъицирована, особенно сзади; самый синус содержит небольшой сгусток крови, легко отделяющийся; в сииусе имеются множественные сухожильные перемычки. С внутренней поверхности dura mater ничего особенного не представляет. Processus falciformis истончен и продырявлен. Твердая мозговая оболочка по sinus longitudinal's сращена с мягкой. Вес мозга 1 537г. Вес левогополушария 634 г,
261
правого — ЫО г. Вес ствола с мозжечком 224 г; вес мозгового ствола 70 г, мозжечка — 153 г. Мягкая мозговая оболочка помутнена, отечна, местами сращена с веществом мозга; сосуды основания мозга ничего особенного не представляют. Мозг вообще полнокровен. В правом полушарии имеется размягчение в передней части бокового желудочка; размягченная масса серовато-желтого цвета. При ближайшем исследовании оказалось, что размягчению подверглась большая часть п. lenticularis, за исключением незначительного переднего его отдела с прилежащей частью белого вещества. Не пораженным остался весь thalamus, равно и corpus caudatum. В спинном мозгу, в грудной части, начиная с шейного утолщения до поясничного, заметно резкое уменьшение размеров переднего рога с левой стороны.
Листки плевры по всей поверхности сращены, в нижней доле правого легкого каверны величиною с голубиное яйцо. Обе почки плотны, узловаты; поверхность разреза усеяна конгломератами туберкулов. Сердце увеличено в объеме, жирно; клапаны нормальны. Печень полнокровна. В почках пассивная гиперемия. Селезенка увеличена в объеме, полнокровна. На слизистой оболочке тонких кишок при переходе их в толстые — отдельные туберкулы. На слизистой же оболочке толстых — поперечные язвы с подрытыми краями.
Микроскопическое исследование спинного мозга обнаружило атрофию клеток переднего рога с левой стороны, особенно заметную на уровне утолщений. Боковые столбы атрофированы и перерождены.
Мы не будем здесь останавливаться на интересе этого случая в смысле оценки фокусного поражения в nucl. lenticularis и прилежащих областей белого вещества полушарий, приведшего сначала к развитию тонической судороги, а затем к развитию паралича движения противоположной половины тела вместе с анестезией мышечного чувства в парализованных членах при сохранении всех других видов кожной чувствительности. Не будем также останавливаться и на резко выраженной в нашем случае церебральной амиотрофии, обусловленной поражением п. lenticularis и последовательной атрофией клеток передних рогов. Но нельзя не обратить здесь внимания на развитие у больного того своеобразного симптома в парализованной левой стороне, который может быть назван pseudomelia paraesthetica и который в данном случае выражался ложным ощущением движения парализованной верхней конечности при полной неподвижности последней.
Заслуживает внимания, что рассматриваемый здесь симптом сопутствовался со стороны чувствительной сферы одной лишь анестезией мышечного чувства, тогда как в ранее описанном мною случае, а равно и в случае приват-доцента П. Останкова, нечувствительность была общею и захватывала все вообще виды чувствительности. С другой стороны, заслуживает внимания, что рассматриваемый симптом обнаружился при строго ограничен ном поражении в области п. lenticularis с прилежащей частью белого вещества.
Уже в первой своей работе я сравнивал испытываемое больным ощущение с тем ложным чувством существования конечности у ампутированных лиц, у которых чувствуемая конечность давно отнята. Аналогия тем более представляется полной, что
262
и в случаях ампутаций ложное ощущение присутствия конечности не исправляется ни сознанием произведенной операции удаления конечности, ни зрением, которое также убеждает в отсутствии ампутированной конечности. Равным образом у ампутированных под влиянием раздражения перерезанных нервов нередко испытывается не только отсутствие удаленной конечности, но иногда и ощущение движения отсутствующей конечности, которое как я мог убедиться в одном из случаев, отличается иногда необычайным упорством и даже мучительностью. Между гем нарушением мышечного чувства при tabes dorsalis, приводящем к обманам в отношении положения конечностей, вышеописанное явление имеет мало общего, так как иллюзии, основанные на анестезии мышечного чувства при tabes, наблюдаются лишь при закрытых глазах и тотчас же исчезают, как только больной открывает свои глаза.
Руководясь вышеприведенным нашим наблюдением, первоначально мною описанным случаем, наблюдением приват-доцента П. Останкова и теми случаями, которые наблюдаются, как последствие ампутаций, мы убеждаемся, что рассматриваемый симптом pseudomelia paraesthetica может обнаруживаться как при церебральных, так и при спинномозговых, а равно и при периферических поражениях, причем, судя по крайней мере по вышеописанному случаю, рассматриваемый симптом должен находиться в теснейшем соотношении с поражением проводников мышечно-суставного чувства, на основании которого создаются ощущения положения различных частей тела и его конечностей.
ОБ ОСОБОМ СГИБАТЕЛЬНОМ РЕФЛЕКСЕ НОЖНЫХ ПАЛЬЦЕВ
В заседании научных собраний клиники душевных и нервных болезней от 22 февраля 1901г. мной был демонстрирован и описан1 в числе нескольких новых рефлексов особый «предплюснопаль-цевой рефлекс, выражающийся легким сгибанием пальцев при постукивании по тыльной поверхности предплюсневых костей и по основанию плюсневых». Этот «сгибательный рефлекс ножных пальцев», как я тогда же отметил, наблюдается лишь при патологическом повышении рефлексов, причем больные, на которых демонстрировался этот рефлекс, представляли собою органическое поражение мозга
В «Neurologisches Zentralblatt», № 5 за 1904, доктор К. Мендель под названием: «Ein Reflex am Fussriicken» описал, очевидно, тот же рефлекс, который мной был описан для патологических случаев, с органическим поражением мозга; но вместе с тем автор указал на то, что рефлекс этот наблюдается и у здоровых лиц, выражаясь здесь не подошвенным, а тыльным сгибанием II—V пальцев, как и в случаях функциональных параличей нижних конечностей; при органических же параличах в одних случаях этот рефлекс может выражаться тыльным сгибанием, в других же подошвенным сгибанием пальцев, причем это последнее видоизменение наблюдается в тех же случаях, в которых обнаруживается и рефлекс Бабинского.
По этому поводу я должен заметить, что разгибательное движение ножных пальцев при постукивании тыльной части стопы наблюдалось также и мной в некоторых случаях у здоровых лиц, хотя и не с абсолютным постоянством, а иногда также и в патологических случаях, но такого рода разгибательное движение пальцев, по моим предположениям, должно было зависеть от механического раздражения m. extensor digit, comm. longus, частью же от непосредственного раздражения подлежащего m. extensor digit, comm. brevis, тогда как сгибательный рефлекс
1 См. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900—190J гг., стр. 44—47.
264
ножных пальцев представляет собою совершенно особый рефлекс, происходящий на счет действия мышц, сгибающих пальцы, и наблюдаемый исключительно только в патологических случаях.
В этом отношении рассматриваемый рефлекс имеет более или менее полную аналогию в описанном мною же сгибательном рефлексе ручных пальцев \ который получается при постукивании молоточком области тыла запястья и основания пястных костей ручной кисти и наблюдается обыкновенно также при патологическом повышении рефлексов (например, при органических гемиплегиях).
Вышеуказанное сообщение доктора Менделя выдвигает, между прочим, важное диагностическое значение сгибательного рефлекса ножных пальцев, которое автором отмечается в двух направлениях: 1) для отличия органических параличей и 2) для определения стороны паралича при коме.
В первом случае существование подошвенного сгибания пальцев при вызывании рефлекса постукиванием говорит, по автору, за органическое поражение, во втором случае, т. е. при коме, то же явление говорит за существование органического паралича на стороне этого явления.
Что касается своих наблюдений, относящихся до описанного мною сгибательного рефлекса ножных пальцев, то я должен заметить, что ни однажды этот рефлекс не был мной отмечен у совершенно здоровых лиц.
При истерической гемиплегии его также не удалось мне отметить ни в одном случае, но он почти с постоянством наблюдался мною при тех из органических параличей (например, при гемиплегиях, при миэлитах и пр.), при которых вобще наблюдается более или менее значительное повышение рефлекторной деятельности.
При этом в резко выраженных случаях описанный мною рефлекс характеризуется не просто лишь подошевенным сгибанием ножных пальцев, но и более или менее ясным растопыриванием пальцев, вследствие их отведения; вызывается же он в таких случаях не только постукиванием с тылу стопы в области предплюсневых и плюсневых костей, но и постукиванием по наружной стороне заднего конца пятой плюсневой кости.
Что касается до отношения сгибательного рефлекса ножных пальцев к рефлексу Бабинского, то на основании своих наблюдений я могу заметить, что почти во всех случаях, где имелся рефлекс Бабинского, я мог наблюдать и вышеуказанный сгибательный рефлекс ножных пальцев; но в круге моих наблюдений имелись случаи с органическим поражением двигатель-
1 В. Бехтерев, О запястно-пальцевом рефлексе, Обозрение психиатрии, 1903; Ueber carpo-metacarpalen Reflex, Neurol. Zbl., 1903, S. 195.
235-
ных проводников, где рефлекс Бабинского отсутствовал или был выражен неясно, тогда как сгибательный рефлекс ножных пальцев существовал налицо.
Таким образом, сгибательный рефлекс ножных пальцев, наряду с рефлексом Бабинского, может служить полезным диагностическим признаком органического поражения двигательных пирамидных путей и притом он может быть использован для доказательства органического поражения двигательных проводников иногда даже в тех случаях, в которых рефлекс Бабинского отсутствует или выражен неясно.
О НАСИЛЬСТВЕННОМ ПЛАЧЕ ПРИ МОЗГОВЫХ ПОРАЖЕНИЯХ
В работе «Насильственный смех и плач при мозговых поражениях» х я обратил внимание на то, что в известных случаях мозговых параличей развивается беспричинный насильственный смех. Тогда же я указал и на случаи мозговых параличей, в которых подобным же образом обнаруживался насильственный плач.
Так как в выражении эмоций, согласно моим экспериментальным исследованиям2, играют роль зрительные бугры и принадлежащие им системы волокон, то очевидно, что такой симптом, как насильственный смех и плач, не может происходить без отношения к его проявлению систем зрительных бугров.
Вслед за мной на случаи мозговых поражений с насильственным смехом и плачем стали обращать внимание и другие авторы (Бриссо, Федоров, Минганзини). Между прочим, Бриссо 3 вскоре опубликовал подобные же случаи, причем подобно мне отнес причину своеобразного симптома к зрительному бугру.
Вышеописанные случаи, как мне кажется, не совсем правильно называть спазмодическим или судорожным смехом, как делает Бриссо и некоторые другие авторы, так как в сущности в этих случаях дело идет о смехе, который, правда, развивается с непреодолимой силой, которого больные удерживать не могут и при котором больные часто даже не могут указать определенно повода к смеху, но тем не менее они всегда определенно утверждают, что им «смешно».
Следовательно, здесь имеется налицо психический импульс, побуждающий к смеху, но нет лишь соответствующей задержки и потому смех в этих случаях развивается при незначительней-
1 В. Бехтерев, Нервные болезни, 1894, в. 1, стр. 49 и след.; см. также: В. Б е х т е р е в, Об отправлении зрительных бугров у животных и человека, Вестник психиатрии, 1885, и Virch. Arch., 1887, Bd. 110 и Arch. f. Psychiatrie, 1897.
8 В. Б e x т е р е в, Об отправлении зрительных бугров у животных и человека, Вестник психиатрии, 1885; Virch. Arch., 1887.
8 Brissaud, Sur le rire et le pleurer spasmodique, Lesons, 1895; см. также. Rf.vue rclentif., 1894, N 2.
267
шем поводе вопреки желанию больного, как навязчивое душевное состояние, подобно навязчивой эмоции.
Но от вышеуказанных состояний навязчивого или насильственного смеха следует, по моему мнению, отличать действительно спазмодический или судорожный смех. Под этим названием следует разуметь такой смех, который развивается без психических поводов, как настоящая судорога. Он действительно наблюдается иногда в более редких случаях эпилепсии в форме ауры при начале приступа. К случаям судорожного смеха следует отнести также не очень давно описанное мной наблюдение Ч
В этом случае дело шло об органическом поражении основных частей мозга с односторонним параличом и развитием односторонних судорог, которые всегда начинались судорожным смехом. Приступам предшествовали каждый раз аура в форме щекотания парализованной левой руки, и самый смех протекал в бессознательной сфере, являясь в настоящем смдсле слова судорожным, как и тоническое напряжение парализованной руки, развивавшееся с постоянством в течение этих приступов. Руководясь тем, что у больной имелась гемианопсия с гемиопической реакцией зрачков, что могло быть объяснено поражением tractus optici или наружного коленчатого тела, я пришел к выводу, что дело идет в этом случае о поражении зрительного бугра, раздражение которого и служило причиной развития спазмоди-ческого смеха.
Особенностью такого судорожного или спазмодического смеха, кроме отсутствия для него соответствующего психического импульса, служит также тот факт, что смех в этом случае сопутствует приступу эпилепсии и развивается в виде его предвестника, безразлично, будет ли дело итти о приступе большой или малой эпилепсии.
Руководясь всем вышесказанным, можно признать, что судорожный или спазмодический смех, о котором здесь идет речь, может быть уподоблен настоящей судороге, тогда как насильственный смех, описанный мною при мозговых параличах, представляет собою явление, психически обусловленное, т. е. наступившее вследствие известных ассоциаций, но развивающееся неудержимо против воли и желания больного, следовательно, может быть уподоблен навязчивым движениям.
Первоначально мной2, а затем и Бриссо3 был высказан взгляд, что такое состояние объясняется устранением задерживающего влияния со стороны произвольно-двигательного пути для facialis, проходящего в переднем отделе внутренней капсулы. Но впоследствии я отказался от этого взгляда, особенно когда мне встретились случаи развития насильственного смеха при
1 В. Бехтерев, Обозрение психиатрии, 1901, стр. 121.
! Тая же.
3 Е г i s s a u d. 1. с.
268
мозговых поражениях без паралича facialis. Такой случай был представлен мною несколько лет назад в одном из научных собраний клиники душевных и нервных болезней 1.
Наряду со случаями насильственного смеха при мозговых поражениях встречаются и случаи насильственного плача при мозговых процессах. На эти случаи насильственного плача я обратил внимание еще в 1894 г. в своей книге «Нервные болезни» (в. 1, стр. 73), где я говорю по поводу этого следующее: «Необходимо заметить, что насильственный или неудержимый плач наблюдается довольно нередко при мозговых поражениях. Мне удавалось его видеть в прогрессивном параличе, при мозговом сифилисе и в случаях мозговых опухолей и при размягчениях. Как известно, насильственный или неудержимый плач встречается нередко и при неврозах, являясь обычным спутником истерии. Но мне случалось видеть насильственный или неудержимый плач как совершенно самостоятельное явление при отсутствии всяких явлений истерии. В этом случае плач, повторявшийся приступами, не сопровождался рыданиями, но приводил всегда к обильному слезоотделению» 2.
Наибольшего интереса в отношении выяснения патогенеза этого своеобразного симптома, как мне кажется, заслуживают случаи гнездовых поражений мозга с развитием насильственного плача. При этом должно иметь в виду, с одной стороны, случаи насильственного плача, обнаруживающегося наряду с существованием насильственного смеха, которые мною уже ранее были описаны3, и затем случаи, в которых при наличности мозгового поражения обнаруживается только насильственный плач без соответственных явлений насильственного смеха.
Такие случаи не часты, но мне встречались уже несколько раз. Одним из типических случаев этого рода может служить, между прочим, следующий случай.
Больной Ш., около 60 лет. Лет 7 тому назад после воспаления легких имел какое-то заболевание, при котором обнаруживалась параличная слабость в конечностях с болями. Со временем больной оправился, но с тех пор у него обнаруживается общая слабость и в особенности слабость ног, которая стала особенно заметно прогрессирвать последние три года.
Перед пасхой 1904 г. больной без видимой причины ослабел в такой степени, что ходить уже не мог, но затем мало-помалу несколько оправился и стал передвигаться, хотя и с трудом. Вместе с развитием слабости ног у больного стал обнаруживаться насильственный плач, появляющийся без всякой причины или при самом незначительном поводе. Достаточно сказать, что во время разговора с врачом больной начинает по многу раз плакать, всхлипывая, не будучи в состоянии удержаться от плача.
1 См. Отчеты научных собраний клиники душевных и нервных бо^ лезней, 26 октября 1900 г.
2 В. Бехтерев, Нервные болезни, в 1. стр. 73.
11 Там же.
269
Когда больной перешел под мое наблюдение, я нашел у него явления склероза артерий, ясно выраженного в лучевых, височных и сонных артериях, резкую глухость сердечных тонов с нечистым оттенком первого-тона. Исследование других органов не дало никаких существенных уклонений. Но у больного имелась параличная слабость нижних конечностей, особенно левой, вследствие чего походка больного представляется паретической, с мелким шагом в виде семенящей походки. По той же причине больной с трудом садится и должен придерживаться руками за что-нибудь, прежде чем сесть. В руках можно также обнаружить некоторую слабость, особенно слева. Складки лица сглажены, язык высовывается прямо. Чуствительность без изменений. Сухожильные рефлексы на нижних конечностях слева повышены. Мочеиспускание ослаблено, на ночь подкладывается утка.
В психическом отношении никаких явлений не обнаруживается, кроме некоторой слабости памяти.
Особенностью больного является неудержимый плач, который появляется без достаточного внешнего повода, иногда при одном виде вновь пришедшего к нему знакомого человека, при первом его слове, притом плач нередко развивается приступами, сменяя один приступ другим, так что правильная беседа с больным становится невоможной. О причине плача больной не дает никаких пояснений, да и не может их дать, так как с каждым вопросом на счет плача последний развивается с такой неудержимой силой, что ни о каком объяснении не может быть и речи. Достаточно больного спросить, почему он плачет, чтобы он начал неудержимо плакать. Когда он успокоится, новый вопрос вызывает тот же неудержимый плач. Лишь иногда больной на вопрос скажет «не знаю» и заплачет.
В обыкновенной домашней обстановке больной также часто и- по самому незначительному поводу плачет. Новое лицо, пришедшее к нему, также возбуждает в нем плач. Всякое радостное событие, а тем более в какой-либо мере печальное, вызывает также неудержимый и продолжительный плач.
Бывают дни, когда больной буквально не в состоянии сказать ни сдова из-за неудержимого плача.
В беседе с больным при веселом его настроении он обнаруживает иногда улыбку и даже смех, но насильственного смеха вовсе не замечается; напротив того, часто начавшаяся улыбка переходит в плач.
С течением времени все явления, обнаружившиеся в физической сфере, стали медленно прогрессировать, мало-помалу у больного развилась столь значительная параличная слабость, что он стал ходить только с поддержкой и вместе с тем голос ослабел в такой степени, что больной мог давать почти шопотом короткие ответы на задаваемые вопросы; насильственный плач продолжался в прежней степени.
После перенесенной пневмонии слабость еще более усилилась, у больного время от времени начали появляться припадки зыбывчивости и недостаточной ориентировки в окружающем, что, впрочем, обычно не продолжалось более нескольких часов и самое большее день-два. В последний период времени у больного развилось апоплектоидное состояние с повышением температуры, после которого слабость достигла такой степени, что больной хотя и мог еще удерживаться на ногах, но почти совершенно не мог ходить, не мог говорить и вместе с тем у него обнаружилось резкое затруднение глотания, чейн-стоксовское дыхание и резко ухудшилась деятельность мочевого пузыря.
Хотя состояние больного с течением времени вновь стало несколько улучшаться, но затем спустя несколько недель у него развилась задержка мочи, присоединился катарр мочевого пузыря, а затем острый пиэлит, от которого он и скончался при высокой температуре в сопорозном состоянии. За последний период болезни до развития сопорозного состояния у больного также обнаруживался неудержимый плач в крайне резко выраженной степени, вследствие чего правильные расспросы об его состоянии представлялись невозможными.
270
Данный случай, в котором имелся насильственный плач без насильственного смеха, представляет собой явления общего прогрессирующего пареза нижних и частью верхних конечностей с преобладанием паретической слабости на левой стороне, — явления, развившиеся на почве обширного склероза мозговых сосудов, повлекшего, очевидно, к гнездовым размягчениям в мозговых полушариях, как это нередко бывает в старческом возрасте.
Но подобные же случаи насильственного плача встречаются и при более ограниченных гнездовых поражениях мозга, как показывает, например, следующий случай, исследование которого было произведено по моей просьбе доктором П. А. Останковым.
Больной 3., 64 лет; себя считает больным около года. 17 августа 1905 г. в 2 часа ночи с ним вдруг сделалось худо, но, оправившись и не потеряв сознания, он лег в постель; через полчаса при позыве к мочеиспусканию он встал, но затем вдруг упал, причем у него скривило рот на сторону и отнялась левая рука и нога. С этих пор больной слег в постель и вылежал около трех месяцев. Дня 3 — 4 после паралича ему плохо повиновался язык. Только спустя три месяца появились движения в ноге, а затем немного и в руке; первое время больной не мог ни стоять, ни ходить, руки висели, как плеть. Впоследствии, кроме полупаралича, у больного еще развилось недержание мочи и стал появляться насильственный или неудержимый плач.
Из анамнестических сведений о больном известно, что отец его умер от мозгового паралича, мать — от водянки; одна из сестер — от чахотки.
В семье нервных и душевнобольных не было. Сифилис отрицает, пил одну меленькую рюмочку водки перед обедом, курил много. Из болезни хворал только в молодости «горячкой», о которой много не помнит и рассказать не умеет.
Status praesens. Зрачки неравномерны, левый зрачок несколько шире правого, световая реакция их выражена хорошо. В лице заметна резкая асимметрия, левая носогубная складка сглажена, левый угол рта опущен; при высовывании языка он немного отклоняется влево. Движения глазных яблок нормальны. Зрение, слух и вкус хороши. Обоняние ослаблено. Левая рука полусогнута и приведена к туловищу; движения в ней ограничены, мышечная сила ослаблена резко. Походка больного паретическая, при ходьбе он приволакивает левую ногу, на одной левой ноге стоит с трудом, мышечная сила в ней ослаблена. При исследовании чувствительности замечается небольшое понижение болевой чувствительности на левой руке, ноге и на левой половине лица.
Коленные рефлексы резко повышены на левой ноге. Слабо выраженный симптом Бабинского. Подошвенный рефлекс получается резко. Со стороны тазовых органов следует отметить запоры и недержание мочи.
Во внутренних органах: гипертрофия сердца (увеличение его границ), диастолический шум, притупление на месте выслушивания аорты. Акцент на a. carotis с правой стороны особенно. Аритмия пульса. Склероз венечных артерий. Полный замедленный пульс.
Что касается насильственного плача, то он у больного появляется уже при простом опросе о его состоянии. Достаточно его спросить о здоровье, чтобы он тотчас же расплакался. Нечего говорить, что один намек на его плач тотчас же приводит к развитию неудержимого плача. На вопрос о причине плача он поясняет, что у него в груди сразу как бы стесняет и приливает к голове, возбуждая наклонность к плачу.
Исследование его физического состояния также обыкновенно приводит к неудержимому плачу. Иногда кажется, что больной плачет без видимой причины, но при расспросах больной обыкновенно указывает тот или иной незначительный повод, например, говорит о своем неизле-
271
чимом состоянии или что-либо подобное. Вообще больной обнаруживает плач почти при каждом обращении к нему с расспросами, иногда в виде многократных приступов и, кроме того, обнаруживает нередко плач и в то время, когда он предоставлен самому себе. Во время представления его мной на лекции слушательницам Женского медицинского института, приступы плача почти не прекращались в течение всего времени, пока больной оставался в аудитории.
Как и в предыдущем случае, здесь дело идет, очевидно, так же]р склерозе мозговых сосудов, но самое поражение должно быть поставлено в связь с местной закупоркой, как надо думать, в области центральных ветвей правой a. fossae Sylvii, чем и объясняется развившийся у больного половинный паралич, к которому затем присоединилось недержание мочи и насильственный плач.
Заслуживает внимания, что как в этом, так и в предыдущем случае насильственный смех сопутствовался недержанием мочи. Этот факт обращает на себя внимание в том отношении, что подкорковые центры мочеиспускания, как показали наши исследования 1, заложены в thalamus opticus, вследствие чего является возможным предположить, что область поражения в подобного рода случаях так или иначе вовлекает в поражение и зрительный бугор. Это же обстоятельство объясняет нам и появление насильственного плача у больных, так как зрительный бугор содержит в себе центры мимических движений 2.
Таким образом, по отношению к патогенезу насильственного плача как симптома мозговых поражений я должен, собственно, повторить то, что сказано мной еще в статье: «Насильственный или неудержимый смех и плач при мозговых поражениях»3.
«Что касается* физиологических условий появления плача, то, без сомнения, они представляют собой много общего с подобными же условиями появления смеха. Плач, возбуждаемый психическими импульсами, без сомнения, обусловливается сочетанной деятельностью мимического центра facialis и центров слезоотделения. Отсюда очевидно, что центрами и проводниками, служащими для импульсов, ведущих к обнаружению плача, являются также зрительные бугры и принадлежащая им система волокон. Таким образом, мы принимаем, что и насильственный плач, встречаемый при мозговых поражениях, имеет в общем туже физиопатологическую основу, как и насильственный смех; следовательно, он обусловливается или устранением задерживающих импульсов или же вызывается раздражением соответствующих центров и проводников».
1 В. Бехтереви Н. Миславский, Центры мочевого пузыря в головном мозгу, Архив психиатрии, 1888, и Neurol. Zbl., 1889.
2 В. Бехтерев, Об отправлении зрительных бугров у животных и человека, Вестник психиатрии, 1885; Virch. Arch., 1887; Основы учения о функциях мозга, в. IV, 1905.
3 Нервные болезни, Казань, 1894, стр. 73.
О РЕФЛЕКТОРНЫХ ЯВЛЕНИЯХ ПРИ ОРГАНИЧЕСКИХ ПАРАЛИЧАХ ЦЕНТРАЛЬНОГО НЕВРОНА И О НОВОМ ТЫЛЬНО-СТОПНОМ СГИБАТЕЛЬНОМ РЕФЛЕКСЕ НОГИ
Как известно, в современной невропатологии было выдвинуто значение некоторых рефлекторных явлений как признаков, дающих возможность отличать органические поражения мозга от так называемых функциональных.
Особенно в этом отношении посчастливилось параличам центрального нейрона, захватывающим нижнюю конечность.
Как верный указатель органического паралича был выявлен прежде всего всем известный клонус стопы, обусловленный пассивным растяжением икроножных мышц при тыльной флексии стопы. Явление это, как было указано мною и некоторыми другими авторами, встречается, хотя и крайне редко, и при истерии \
Но, как недавно было доказано исследованиями в заведывае-мой мной клинике Женского медицинского института (М. П. Никитин), имеется возможность отличить путем графической записи «органический» клонус как более или менее равномерный от истерического, не отличающегося такой равномерностью.
Явление другого порядка, но также относящееся к рефлекторному эффекту пассивного мышечного растяжения, было уже давно описано мной при параличах центрального нейрона на верхних конечностях, причем оно, как показали мои же наблюдения, обнаруживается при одинаковых условиях и на нижних конечностях 2.
Явление это заключается в том, что если парализованную верхнюю конечность сгибать и разгибать, то при известной быстроте этого разгибания испытывается препятствие на определенном пункте (приблизительно на середине расстояния между положением предплечья под прямым углом и полным его разгибанием), после чего разгибание продолжается свободно. Иногда то же явление можно обнаружить при сгибании предплечья, ? в более редких случаях и при супинации. Аналогичное явление со стороны нижней конечности состоит в том, что если парализо-
1 См. В. Бехтерев, Невропатологические и психиатрические наблюдения и исследования, СПБ, 1910, вып. 1, стр. 237. 'Там же.
18 в. М. Bex-rep?!}
273
ванную нижнюю конечность быстро согнуть в коленном суставе, то она не сгибается сразу, а обнаруживает при известном угле сгибания рефлекторное противодействие, после которого сгибание продолжается уже беспрепятственно.
То же нередко обнаруживается и при пассивном разгибании ноги в коленном суставе, причем как при сгибании, так и при разгибании рефлекторное противодействие обычно обнаруживается всегда в определенном пункте сгибания и разгибания ноги, большей частью при положении ноги, образующей в коленном сгибе угол больше прямого. При этом вышеуказанное препятствие обнаруживается лишь при том условии, когда самое сгибание или разгибание производится быстро; при медленном же сгибании и разгибании это явление не обнаруживается совсем.
Затем, как известно, получил особое значение всем известный ныне рефлекс Бабинского, получающийся при механическом раздражении подошвы и состоящий в тыльном сгибании большого пальца ноги, сопровождаемом нередко разгибанием и растопыриванием других пальцев, тогда как в нормальном состоянии, за исключением младенческого возраста и вообще возраста первого детства, подошвенное раздражение вызывает обыкновенно подошвенное же сгибание пальцев стопы.
Как видоизменение рефлекса Бабинского, состоящее в вызывании его раздражением других областей, следует рассматривать рефлекс Оппенгейма, заключающийся в том же разгибательном движении большого пальца, но вызываемый штриховым раздражением рукояткой молотка наружных покровов голени.
Аналогичное значение должен иметь и рефлекс Шеффера, представляющий то же явление со стороны большого пальца ноги, но вызываемое простым сдавливанием ахиллова сухожилия.
Со своей стороны я могу указать еще на один из способов вызывания разгибательного движения большого пальца, а иногда и других пальцев ноги, состоящий в сильном сдавливании мышц голени и бедра, которое я описал еще несколько лет назад. Явление, о котором идет речь, состоит в том, что, если захватить рукой толщу икроножных мышц у больного с поражением центрального нейрона и затем сильно сдавить их, то получается раз-гибательное движение пальцев ноги, особенно большого. Аналогичное же рефлекторное разгибание большого пальца получается иногда и при сдавливании мышц бедра.
Совершенно другого порядка явление, наблюдаемое также при параличах центрального нейрона, представляет описанный мной сгибательный рефлекс малых пальцев ноги, вызываемый механическим ударом молотка по наружной части тыла стопы *.
1 К. Мендель, описавший этот же рефлекс несколько лет позднее меня, признавал его извращением нормального рефлекса, получающегося при постукивании тыла стопы и состоящего в разгибательном движении пальцев. Однако с тыла стопы у нормальных лиц можно получить.
L74
Тот же сгибательный пальцевой рефлекс может быть вызван иногда и при механическом раздражении ударом молотка с других частей стопы, например, с пятки или с наружного края стопы
II ПОДОШВЫ.
В нашей же клинике (д-р Жуковский) было выяснено, что вышеописанный рефлекс с особенным постоянством вызывается ударом по средней части подошвы стопы, где он иногда вызывается с большей силой, нежели с тыла стопы 1.
Другое явление, которое мной было описано в «Образовании психиатрии» и в «Neurologisches Zentralblatt» несколько лет тому назад при органических параличах центрального нейрона, представляет собой общий оборонительный рефлекс ноги, вызываемый быстрым подошвенным сгибанием ее пальцев и стопы 2.
В резко выраженных случаях тотчас вслед за быстрым подошвенным сгибанием пальцев и стопы происходит обратное тыльное разгибание пальцев и стопы и последовательное сгибание голени и бедра иногда с небольшим отклонением его кнаружи. В зависимости, однако, от меньшей степени выраженности этого явления и от меньшей силы и быстроты производимого подошвенного сгибания пальцев и стопы, дело может ограничиваться в отдельных случаях только тыльным разгибанием пальцев и стопы или даже одного большого пальца и не сопровождаться дальнейшим эффектом в виде сгибания голени и бедра.
Это описанное мной явление вошло уже в такие руководства по неврологии, как Левандовского, хотя оно истолковывается им почему-то не как рефлекс, а как совместное движение, что, на мой взгляд, неправильно. Затем недавно это явление было доложено Фуа в Парижском неврологическом обществе и затем в Казанском обществе невропатологов и психиатров3, хотя в нашей клинике это явление, как уже давно изученное и описанное мною в литературе, исследуется постоянно у больных и уже в течение многих лет служит для установления диагноза параличей центрального двигательного нейрона.
только разгибательное движение отдельных пальцев (одного или двух) при ударе по соответствующему промежутку плюсневых костей, обусловливаемое, очевидно, непосредственным раздражением подлежащих мышц, вследствие чего я не могу согласиться с мнением Менделя.
1 Аналогично сгибательному рефлексу пальцев стопы мной был списан на кисти руки запястно-пальцевой рефлекс, обнаруживающийся также при параличах центрального нейрона и выражающийся сгибательным движением пальцев руки при ударе молоточком по тылу запястья, особенно ближе к наружной его стороне. И этот рефлекс, который при резком повышении рефлексов может быть вызван также и с других частей кисти руки и, между прочим, с нижнего конца локтевой кости, как оказывается, может быть вызван с особенной легкостью также и ударом молоточка по средней части ладонной поверхности руки.
2 См. В. Бехтерев, Обозрение психиатрии, 1905, стр. 571.
3 В е с е л и т с к и и, Обозрение психиатрии, 19! 1.
18* 275
Заслуживает внимания, что в резко выраженных случаях достаточно быстро подогнуть большой палец стопы, чтобы он тотчас же разогнулся в тыльном направлении.
Очевидно, здесь дело идет о явлении, аналогичном предыдущему, но как бы частичном, а не столь общего характера, как при вышеуказанном приеме подошвенного сгибания пальцев стопы.
В последнее время я имел возможность отметить еще одно явление, которое может служить достоверным признаком органического поражения центрального двигательного нейрона и которое в то же время представляется явлением крайне своеобразным. Оно состоит в том, что если мы станем ударять молоточком по тылу стопы, безразлично в какой именно области, то обычно после нескольких ударов стопа обнаруживает тыльное сгибание, причем пальцы, особенно большой, разгибаются и происходит сгибательное движение всей ноги в коленном суставе.
Речь идет, таким образом, о совершенно своеобразном костном рефлексе, развивающемся под влиянием многократного постукивания по костям тыла стопы как результат суммирования их раздражения.
Мне кажется, что описанный мною же несколько лет тому назад костный рефлекс, получающийся у больных с параличом центрального неврона при многократном постукивании молоточком по передней поверхности большой берцовой кости и состоящий в сгибательном движении ноги в коленном, бедренном и голеностопном сочленении, представляет собою аналогичное с вышеуказанным явлением.
Надо, впрочем, заметить, что описываемый ныне сгибатель-ный рефлекс ноги не представляется исключительно костным, так как в тех случаях, где он вызывается постукиванием молоточка по тылу стопы, нередко он может быть вызван одинаково, и при уколах в тыл и другие части стопы и вообще при нанесении болевых раздражений, причем обращает на себя внимание, что в этом случае рефлекс не носит характера движения, отстраняющего стопу от колющего орудия, а, напротив того, как бы направляющего стопу на колющее орудие.
Нет надобности входить здесь в сравнительную оценку каждого из вышеуказанных симптомов, служащих признаками параличей центрального неврона. Необходимо лишь иметь в виду, что ни один из этих признаков, не исключая клонуса и рефлекса Бабинского, не составляет абсолютно постоянного признака при параличах центрального неврона.
Случается, наоборот, чаще всего так, что налицо имеется один или два из вышеуказанных признаков, другие же отсутствуют. Неоднократно отмечалось, например, отсутствие рефлекса Бабинского при существовании сгибательпого пальцевого рефлекса стопы и, наоборот, может существовать рефлекс Бабинского и отсутствовать сгибательный пальцевой рефлекс.
276
Отсюда понятно, почему нельзя довольствоваться в диагно» стических целях одним каким-либо признаком, а приходится принимать во внимание все вообще признаки, которые могут иметь в каждом отдельном случае известное значение.
Что касается вновь описываемого признака, состоящего в сгибательном движении нижней конечности при постукивании молоточком по тылу стопы, то он принадлежит, вообще говоря, к числу более или менее постоянных признаков параличей центрального нейрона, обнаруживаясь как при миэлитах и других поражениях спинного мозга выше поясничного утолщения, так и при поражениях головного мозга (размягчение, кровоизлияние и пр.), прерывающих двигательные пути.
При этом мне встречались случаи, когда рассматриваемый рефлекс обнаруживался в параличах центрального нейрона при отсутствии рефлекса Бабинского и даже клонуса стопы, а равно и описанного мною сгибательного пальцевого рефлекса. Это убеждает меня в том, что вновь описываемый рефлекс не лишен диагностического значения в подходящих случаях.
О РЕФЛЕКТОРНОЙ ЭПИЛЕПСИИ ПОД ВЛИЯНИЕМ ЗВУКОВЫХ РАЗДРАЖЕНИЙ
Как известно, давно установлено существование так называемой рефлекторной эпилепсии, обусловленной болезненными рубцами травматического происхождения. В этом отношении литература обладает немалым числом убедительных случаев, в которых не только развитие падучих приступов началось после травматического повреждения, обусловливавшего развитие болезненного рубца, не только самый приступ начинался чувством особого раздражения в области этого рубца в форме ауры, после чего судорога, начавшись с того органа, который пострадал при травме, постепенно распространяется на другие части тела и, наконец, на все тело, сопровождаясь в конце потерей сознания, но и тем, что с удалением раздражающего рубца оперативным путем прекращались и судорожные припадки.
Имея в виду, что в иных случаях не менее раздражающие периферические повреждения не вызывают совершенно аналогичных явлений, необходимо признать, что в этих случаях дело идет, очевидно, об особой возбудимости корковых центров, находящей себе объяснение в причинах общего характера.
Недавно в русской литературе было обращено внимание на то, что и зрительные раздражения в известных случаях могут вызывать развитие эпилептических припадков, причем моментом, возбуждающим припадки, являлось быстрое смещение света и тени, приводившее к утомлению органа зрения, за которым следовало развитие бессознательного состояния и судорог. •
Заслуживает, далее, внимания вопрос с развитии эпилептических припадков вследствие раздражения органа слуха специфическими раздражениями. В своей практике я встретился уже в двух случаях с развитием падучих приступов в связи с телефонированием. В этих случаях уже дребезжание телефонного звонка было достаточно для возбуждения эпилептического приступа.
Но несомненно, что и музыка сама по себе способна вызывать в некоторых случаях развитие эпилептических приступов. В этом отношении в литературе мы имеем указание на развитие падучих приступов под влиянием музыки в описании профессора И. П. Мержеевскогс. Мне встретилось уже два случая, в ко-
:-/8
торых эпилептические приступы вызывались музыкой. В виду особого интереса такого рода случаев я могу привести здесь описание одного очень характерного случая этого рода, который тем более поучителен, что относится к интеллигентному лицу, известному литератору Б. Никонову. В этом случае развитие болезненного состояния подвергнуто им же самим подробному анализу в рукописи, любезно предоставленной в мое распоряжение, что еще более увеличивает ценность этого наблюдения.
Ввиду особого интереса этого случая я приведу самую рукопись полностью, без малейших сокращений.
Она озаглавлена им:
Страх музыки
«Я был гимназистом второго или третьего класса, когда со мной впервые произошло то неприятное ощущение от музыки, которое устрашает и угнетает меня ныне.
Я очень люблю музыку и часто бывал в опере и в концертах. И вот, когда однажды я был на представлении «Гугенотов», и когда на сцене шел второй акт, в котором очень долго поет в начале акта королева Маргарита, я почувствовал странное и испугавшее меня ощущение какого-то внезапного удара, или судорожного сжатия в горле и одновременно с тем подергивания в левом глазу. Это ощущение было весьма непродолжительно и по существу ничего неприятного и страшного собою не представляло, но оно испугало меня как нечто никогда дотоле со мной иебывавшее и необычное.
Мне казалось, что оно произошло от продолжительного и напряженного созерцания блестяще одетой королевы, ярко освещенной театральным солнцем. Поэтому я решил совсем не смотреть на сцену и чем-нибудь отвлечь свое внимание от сценического действия и музыки. Я стал рассматривать ряды лож и кресел, прислушиваться к посторонним звукам, рассматривать свои карманные часы. Поуспокоившись, я опять занялся созерцанием оперы —и на этот раз совершенно благополучно. Испугавшее меня подергивание в глазу и удар в горле уже не повторялись.
После того я не запомню в течение очень долгого периода времени, чтобы со мной случались подобные явления на оперных представлениях. Может быть, это и бывало, но не оставило точного воспоминания. Во всяком случае, до зрелого возраста (при близительно лет до 20—22) я мог бы насчитать очень большое число спектаклей и концертов, на которых я присутствовал очень благополучно. Но я должен отметить следующее обстоятельство: совершенно благополучно проходили те сеансы музыки, на которых я присутствовал не один, но с кем-либо из знакомых Мне лиц, с которыми я мог во время музыки перекинуться словом и таким образом отвлечь свое внимание от непрерывного гула
279
музыки. А такие спектакли и концерты, на которых я присутствовал совершенно один —насколько я могу помнить —даже в то сравнительно благополучное для меня время (т. е. до 20 — 22 лет) иной раз сопровождались пугавшими меня ощущениями, не в такой степени, однако, чтобы я серьезно обращал на эти ощущения внимание.
Тогда я не боялся слушать оперную и вообще массовую громогласную музыку. Но вот наступила эпоха, когда я стал бояться музыки.
Это было, когда я был студентом, уже оканчивавшим курс. Несмотря на то, что неприятные ощущения к этой поре стали учащаться, и неизбежно случались, когда я присутствовал на музыке один, т. е. без сопровождения кого-либо из близких лиц, я храбро посещал спектакли и ничуть не придавал рокового значения упомянутым ощущениям. Они обычно приходили ко мне в середине спектакля (конец второго или начало третьего акта). Я, правда, приходил в волнение от загадочных подергиваний глаза и ударов в горле, покрывался от волнений испариной и даже, кажется, дрожал, но принятыми мерами (отвлечение внимания, затыкание ушей, временный уход из залы) успокаивал себя, и после пережитого в середине спектакля кризиса ощущения проходили, и я благополучно и с удовольствием досиживал спектакль до конца и стремился затем на новые спектакли.
Но нижеследующий случай внушил мне настоящий страх к музыке, после чего я уже стал избегать ее.
В описываемый период времени я однажды присутствовал (в одиночестве) на представлении «Пророка» в Мариинском театре. Я уже был на «Пророке» за несколько дней до этого спектакля и так как в тот первый раз я был вместе с сестрой, то ничего удручающего со мной не произошло. Но не то ожидало меня на этом втором представлении «Пророка». Я сидел (собственно, стоял, чтобы лучше видеть сцену) в боковой галле-рее на самом верху. Первые акты прошли благополучно. Начался балет (кажется, в третьем акте), изображавший катанье на коньках. Я уже замечал до этого случая, что балетное мелькание пестрых фигур в связи с ритмически-однообразной музыкой чаще всего вызывало у меня страшное ощущение... То же случилось и в этот раз: едва только замелькали пестрые танцовщики, катаясь из угла в угол по сцене, как у меня задергало левый глаз частыми подергиваниями. Горло стала схватывать судорога. Я пришел в волнение, заткнул уши и зажмурил с усилием левый глаз, но удары усиливались. И вдруг я почувствовал, что теряю сознание. Подергивание глаза и удары в горло, все усиливаясь, перешли в такое ощущение, как будто кто-то изо всей силы ударял меня тяжелым предметом по темени. Я погрузился в тьму и с мыслью, что это наступил мой конец, утратил сознание.
280
Я очнулся в коридоре. Меня вели под руки и давали мне пить. В голове я ощущал резкую боль — я ушибся при падении. Зернувшись к жизни, я не сразу сообразил, что такое со мной произошло... Но когда я отлежался в приемной у театрального врача (к сожалению, я совершенно не помню, о чем он расспрашивал меня и расспрашивал ли), я выразил желание итти в зал дослушивать оперу, чему врач весьма энергически воспротивился. Страха к музыке у меня в те минуты не было, быть может, отчасти потому, что ко мне еще не вполне вернулось ясное понимание всего происходящего. Я только потом —спустя несколько дней, вспомнил свое «умирание» и погружение в тьму, а в те минуты просто недоумевал, каким образом я очутился в коридоре и у врача, и мне хотелось использовать до конца свое пребывание в театре.
Как бы то ни было, однако, с этой поры у меня возник уже настоящий страх к музыке. И хотя я еще несколько раз и после того бывал в опере и на концертах, но уже с большой опаской, и едва только начинались роковые подергивания глаза, я убегал из залы, а иногда и совсем покидал спектакль. И мало-по-малу пришел к заключению, что мне совершенно не следует бывать в опере и слушать музыку. Обмороки с той поры со мной уже не случались, но, очевидно, только потому, что я старался не доводить себя до состояния, близкого к обмороку. Если я и бывал в опере, то не иначе, как с кем-нибудь из знакомых и преимущественно в ложе, откуда было удобнее убегать в критические моменты начавшихся подергиваний.
Страх к музыке у меня рос по мере того, как росли неприятные ощущения от нее. Вначале я боялся громкой и близкой музыки. Но потом оказалось, что я имею основание бояться также музыки далекой и тихой: она так же дурно действовала на меня, как и громогласная музыка. Наконец, временами на меня дурно действует даже обыкновенная игра на рояли, и даже в том случае, когда игра звучит за стеной в соседней квартире. А однажды у меня начались подергивания и удары в горле от простой гармоники, на которой играл мальчишка на пароходе, и притом вскоре после того, как я услышал эту игру.
Страх, который охватывает меня, когда я слышу музыку, сопровождается (иногда) сердцебиением и испариной. Если мне приходится встречаться на улице с полковой музыкой, то я проявляю картину полного малодушества: бегу от нее куда глаза глядят —в боковую улицу, в ворота чужого дома или в подвернувшийся магазин, где мне приходится совершать по этому случаю какую-нибудь ненужную покупку. В оперу и на концерты (как общее правило) я, конечно, не хожу теперь, но часто вижу сон, будто мне почему-то приходится сидеть в Мариинском театре, и сон этот мне кажется довольно страшным. Недавно мне пришлось наяву побывать в этом театре на оперном представлении, и хотя действительность оказалась несколько менее страш-
28!
ною, чем сон, но страх все-таки не покидал меня все время, пока я присутствовал на спектакле. Впрочем, понятие «присутствовал» в данном случае уместнее заменить противоположным понятием— «отсутствовал»: я часто уходил из залы, а когда сидел в зале, то тщательно затыкал уши, так что звуки воспринимались мною довольно слабо.
Позволю себе сделать кое-какие выводы касательно моего страха музыки.
На меня действует преимущественно непрерывная и притом в достаточной степени продолжительная по времени музыка. Неприятные ощущения никогда не возникают сразу, одновременно с восприятием первых музыкальных звуков. Необходимо привыкнуть к струящейся звуковой волне, чтобы слух освоился с нею, и тогда упомянутые ощущения неизбежны. Теперь, когда я испытываю страх перед музыкой и когда я начинаю волноваться уже в первые же мгновения после того, как услышу музыку, этот предварительный период «привыкания» чрезвычайно сокращается. Томительное состояние страха и желания провалиться сквозь землю, чтобы только спастись от звуков, приближают ко мне роковой момент. Если музыка сопровождается частыми перерывами, то эти перерывы несколько успокаивают меня и позволяют мне кое-как дослушать пьесу до конца. И чем продолжительнее и чаще перерывы, тем легче ознакомиться мне с музыкой без роковых для меня последствий. Но всего убийственнее зато для меня так называемый «звуковой фон», принятый в современных операх (особенно у Вагнера) — непрерывное течение звуков, являющееся как бы канвой, на которой рисуются различные индивидуальные мелодии и музыкально-драматические сцены. От этого фона нет возможности спастись, он проникает все существо и держит меня в настоящей кабале, единственное спасение от которой —стремительное бегство куда глаза глядят.
Что касается характера звука в зависимости от того или иного инструмента, то я должен сказать, что всего убийственнее на меня действует музыка громкая, какая бы она ни была по свойству самого музыкального инструмента. Если же принять за норму среднюю громкость звука, то я хуже переношу духовые инструменты и несколько лучше струнные, причем опять-таки приходится сделать исключение для скрипок, которые для меня несносны своим зудением и назойливостью своего звука. Рояль и арфа наиболее для меня переносимы. Пение слушаю легче, чем музыку.
-По своему характеру, музыка действует на меня приблизительно одинаково, какая бы она ни была: бравурная или элегическая. Вся суть в силе звука и в его продолжительности. Можно, впрочем, сделать еще маленькое замечание касательно темпов музыки: медлительный темп для меня переносимее, чем темпы быстрые (танцовальные) и притом прерывистые. На-
282
сколько вредно действует на меня непрерывность самого звука и насколько облегчается для меня слушание музыки, разделенной известными паузами молчания и отдыха, настолько легче для меня продолжительный и ровный темп музыки и тяжелее темп прерывистый и беспокойный, обусловленный частыми интервалами, скачками и вибрацией звука. Маршеобразная и танцевальная музыка —самая непереносимая и страшная для меня.
Я считаю совершенно неправильными предположения, будто я обязан своими ощущениями тому душевному волнению, которое производит на меня музыка сама по себе или в связи с драматическим сценическим действием (в опере). Наоборот, если что иногда и спасает меня и позволяет мне кое-как слушать музыку, то именно душевная приподнятость от качества и сюжета музыки. В тех редких случаях, когда мне кое-как удается побороть панический страх пред музыкой и заставить себя слушать ее, и если в таких случаях музыка оказывается приятной для моего слуха или вызывает во мне известное волнение чувства, то я слушаю ее довольно безнаказанно. Если же музыка не будит во мне никакого чувства и оставляет меня совершенно пассивно плавать в ее волнах, то я неизбежно и в весьма непродолжительном времени испытаю свои тягостные ощущения. Эту пассивность слушания, т. е. отсутствие сочувствия к музыке, не следует смешивать с невнимательным отношением к музыке, т. е. с неслушанием ее, с отвлечением от нее. Но я должен оговориться: воздействие музыки на мое чувство и ее приятность для моего слуха вовсе не спасают меня от тягостных ощущений. Они только несколько отсрочивают их.
Было бы крайне желательно поднять вопрос об ограничении пользования музыкой когда попало и где попало. Практикующееся в нашем обществе неумеренное пользование музыкой приводит к тому, что таким лицам, как я, от музыки, в сущности, нет нигде спасенья. Мы слушаем ее тогда, когда не хотим слушать и боимся слушать. Звуки музыки насильно вторгаются r самые сокровенные углы нашего жилища и всюду настигают нас От музыки невозможно спастись, как мы спасаемся от неприятных явлений искусств другого порядка: живописи, поэзии и т. п. Излишне музыкальные соседи ввергают нас в кабалу своему искусству каждое мгновение —иногда даже глубокой ночью. Это крайне тяжело и антиобщественно...».
К сделанному описанию считаю необходимым добавить, что дело идет в данном случае о лице около сорока лет, с наследственным расположением (отец умер от удара), с детства отличавшемся известной нервностью, но в остальном человеком совершенно здоровым, удовлетворительного сложения и довольно упитанным. Его страх к музыке вторичного происхождения и обусловлен, как видно из описания, исключительно наступающим каждый раз появлением тягостных ощущений, как бы удара
283
в голову, и появлением судороги в левом глазу и в горле, являющейся в форме предвестника эпилептического приступа.
Надо заметить, что эпилептический приступ с больным был не единственный, но он всегда являлся последствием слушания музыки, которую больной вообще чрезмерно любит, будучи человеком с хорошим музыкальным слухом.
В последнее время больной начал лечиться, принимая, по моему совету, предложенную мною смесь из адониса, брома и кодеина с содой, и оказалось, что под влиянием этого лечения он освободился как от эпилептических приступов, вызываемых музыкой, так и от страха перед последней. В последнее время он сделался даже завсегдатаем оперных театров, выполняя обязанности музыкального критика.
Заслуживает внимания, что появлению первого эпилептического припадка у больного содействовало «балетное мелькание пестрых фигур» в связи с ритмически однообразной музыкой. Стало быть зрительное мелькание, как агент, играющий роль вообще в развитии эпилептических припадков у некоторых лиц, не остался без значения и в нашем случае. Но во всяком случае наиболее существенную роль в развитии припадков, как видно из предыдущего описания, играла в нашем случае музыка.
Заслуживает внимания, что в данном случае дело шло об эпилептических приступах, вызывавшихся музыкальным воздействием у лица, которое раньше никогда эпилептическими приступами не страдало.
Что касается характера музыки, то на больного больше всего действует громкая музыка, особенно духовая, а также скрипка своим протяжным звуком. Темп быстрый и прерывистый, вообще беспокойный, с частыми интервалами, скачками и вибрацией звуков действует вообще хуже, чем ровный и спокойный темп. В особенности же неблагоприятно действует на больного музыка с темпом маршей и танцев и прямо убийственно музыкальный фон или непрерывное течение звуков в виде звуковой канвы для мелодий, как это принято в современных операх.
После всего сказанного ясно, что мы имеем здесь дело с рефлекторной эпилепсией под влиянием звукового раздражения, как в других случаях мы имеем рефлекторную же эпилепсию под влиянием светового и кожного раздражений.
Что касается сущности самого действия звуковых, в частности музыкальных, раздражений на развитие эпилептических приступов, то оно должно быть поставлено в связь с влиянием слухового раздражения на состояние мозгового кровообращения, что может быть доказано и опытным путем.
О ДВИГАТЕЛЬНОМ И ЧУВСТВИТЕЛЬНОМ ФЕНОМЕНЕ ЛОКТЕВОГО НЕРВА1
Повышенная механическая возбудимость нервных стволов до сих пор мало была использована при нервных болезнях в целях диагностики. В этом отношении можно указать на старые исследования Шарко о нервно-мышечной возбудимости в глубоких степенях гипноза у истеричных, причем те демонстрации, которые мне удалось видеть у самого проф. Шарко в Париже в начале 80-х годов, были в сущности явлениями повышенной мышечной возбудимости к механическим раздражениям. Еще раньше был отмечен Труссо при тетании симптом нервно-мышечной возбудимости при общем сжатии нервных стволов и мышц в нижней плечевой части руки, в результате которого получается напряжение в мышцах кисти и пальцев (рука акушера). Затем при тетании же была отмечена повышенная возбудимость ствола лицевого нерва (Хвостеком) наряду с повышенной нервно-мышечной гальванической возбудимостью, отмеченной Эрбом. Наконец, мною при той же тетании была отмечена нарастающая реакция, или реакция возбуждения, получающаяся как при многократно повторяющемся замыкании гальванического тока на мышцах и нервах, так и при повторном механическом раздражении ударом молоточка, производимым по мышцам и нервным стволам.
В менее выраженных случаях тетании в моей клинике была констатирована и неполная реакция возбуждения (доктор М. С. Добротворский), состоящая в том, что реакция при последовательном ряде замыканий током постепенно нарастает в силе, после чего почти внезапно ослабевает и затем вновь начинает нарастать, но через некоторое время вновь ослабевает, после чего опять начинает нарастать и т. д. То же самое я мог констатировать в подобных случаях и притом не только в отношении электрической, но и в отношении механической реакции мышц. Последняя, подобно первой, нарастая при последовательном ряде ударов по нервам и мышцам, затем быстро ослабевает, после чего вновь начинает нарастать до известного предела и т. д. Такую
1 Доложено в заседании врачей нервной клиники Института медицинских знаний, 1924.
285
реакцию я предложил бы назвать уступающей реакцией возбуждения или, мол-сет быть, еще лучше, «лестничной реакцией».
Если мы отметим затем особую чувствительность нервных стволов при невритах к давлению и, наоборот, отмеченную доктором Бернацким нечувствительность локтевого нерва при его поколачивании, наблюдаемую при спинной сухотке и описанный мною еще более ранний симптом спинной сухотки в виде безболезненности п. poplitci и икроножных мышц при их сдавлении1, то этим почти исчерпываются паши сведения о механической возбудимости нервных стволов.
За несколько последних лет в целом ряде общих неврозов и других случаев нервных заболеваний, характеризующихся спазмофильными проявлениями, обусловленными нарушением функции желез внутренней секреции с преобладанием, повиди-мому, гипофункции околощитовидных желез, я убедился в существовании очень характерного феномена, относящегося также к повышенной возбудимости нервных стволов. Если согнуть предплечье верхней конечности до прямого угла с плечевой частью руки, то, став кнаружи от руки больного, мы легко нащупаем указательным пальцем желобок плечевой кости при локтевом сочленении, в котором лежит локтевой нерв. Если затем пальцы своей руки мы поставим непосредственно на желобке и будем прокатывать под указательным пальцем легко ощупываемый при таких условиях локтевой нерв, зацепляя его, наподобие струны, ногтем своего пальца, то каждый раз, когда локтевой нерв будет проскальзывать, как шнурок, под указательным пальцем, мы получим более или менее резко выраженное сгибателыюе двил<ение пальцев руки, особенно IV и V, а в более слабой мере проявляющееся и в других пальцах кисти, включая и большой палец; в случае же более резко выраженной спазмофильности молеет появляться далее легкое сгибание и всей кисти, а иногда, при особенно повышенной возбудимости, рефлекс обобщается на вышележащие области в виде общего вздрагивания руки, распространяясь иногда и на другую сторону. Этот сгибательный двигательный феномен, наблюдаемый в ряде случаев то в большей, то в меньшей степени, сопровождается более или менее резким ощущением пробегания как бы электрического тока, ползания мурашек и онемения, а иногда и болевой реакцией той или другой степени. При этом чувствительный эффект в виде тока, мурашек или онемения распространяется вдоль предплечья до пальцев кисти включительно, а в более редких случаях идет и вверх в направлении плеча.
Вышеуказанное явление с характером двигательного и чувствительного ' компонента можно вызывать описанным путем любое число раз, причем там, где это явление резко выражено, можно иногда видеть, как вместе с многократным вызыванием
1 ССозренье ьсп.м'^трш!, I'ju-i. Л:< Z, и Neurol. Zbl., 1915, № 21, S. 978.
:.i>6
данного феномена двигательный эффект несколько нарастает до той или иной степени, наподобие описанной мною в свое время при тетании механической и гальванической реакции возбуждения1.
Не может подлежать сомнению, что вышеуказанное явление не исключительно представляет собою механическое раздражение локтевого нерва, но также является и эффектом рефлекторного характера. Это очевидно из того, что в случаях с слабо выраженным явлением мы имеем только эффект сгибания двух последних пальцев руки, тогда как в случаях с более резко выраженным явлением мы имеем не только сгибание двух последних пальцев, но и сгибание других пальцев руки и в том числе большого пальца, и даже легкое сгибание кисти, а иногда и обобщение рефлекса на всю руку и на другую сторону. Эти явления, таким образом, должны быть поставлены насчет участия в раздражении срединного и других нервов, очевидно, путем рефлекса через спинной мозг с распространением его в более редких случаях даже на другую сторону. Наконец, в особо редких случаях я наблюдал извращенный феномен, состоящий при механическом раздражении локтевого нерва в разгибании пальцев руки, зависящем, как известно, от возбуждения лучевого нерва, а иногда и своеобразное перебирание пальцев руки.
Из вышеизложенного очевидно, что описываемый мною «феномен» не является результатом одного механического раздражения локтевого нерва, а является симптомом смешанного характера в смысле прямого механического раздражения нерва и рефлекторного эффекта, осуществляемого через спинной мозг. Что вышеописанный феномен не обусловлен только передачей возбуждения через спинной мозг, это, мне кажется, вытекает из того, что при более слабо выраженной спазмофильности сгиба-тельный эффект может ограничиваться только сгибанием двух последних пальцев. С другой стороны, и чувствительная реакция во многих случаях сопровождается обыкновенно ощущением, идущим от места нажатия к периферии, и только в более резких случаях мы имеем своеобразное ощущение, пробегающее и в центральном направлении по плечевой части руки. Это и заставляет нас склониться к мысли, что в указанном феномене мы имеем явление смешанного характера, т. е. как непосредственное механическое раздражение локтевого нерва, так и рефлекторную передачу возбуждения от места раздражения через спинной мозг.
Во многих сотнях исследованных мною случаев нервнобольных в своей практике и затем у многих больных, исследованных позднее в патолого-рефлексологическом институте совместно с докторами Гаккель и Ратнер, вышеуказанное явление по степени своего развития оказывалось неодинаковым в различных
1 В. Бехтерев, Тетания, Неврологический вестник, т. II, в. 2; Die Tetanie, Dtsch. Zschr. f. Nervenheilk., Bd. VI.
287
случаях. В ряде их двигательного эффекта при вышеуказанных условиях вовсе не наблюдается, несмотря на то, что испытуемые в этих случаях заявляли о движении мурашек или онемении, распространяющемся по ходу нерва, а иногда и болевом эффекте, на месте раздражения. В других же случаях двигательный эффект был выражен, хотя и вполне ясно, но все же едва намечен в IV и V пальцах и часто в большом, а в третьих случаях он выявляется с необычайной резкостью и обнаруживает иногда еще, при многократном повторении опыта, ясную реакцию возбуждения, т. е. постепенного возрастания двигательного эффекта до тою или другого предела1. Последних лиц мы причисляем к категории резко спазмофильных, первых к категории неспазмо-фильных, остальные занимают между теми и другими промежуточную группу, являясь слабо спазмофилы ыми.
Необходимо здесь же отметить, что спазмофильность представляет собою явление функционального характера, а потому она не может отличаться каким-либо особым постоянством и может характеризовать временное состояние того или другого лица. При вызывании феномена локтевого нерва вышеописанным •способом эффект нередко представляется неодинаковым на той и другой стороне, что должно быть поставлено на счет главным образом неодинаковой возбудимости нерва той и другой стороны. В какой мере неравномерность двигательного эффекта на той и другой стороне зависит от анатомических причин, остается еще выяснить.
Вызывание феномена локтевого нерва чаще всего не сопровождается болезненной восприимчивостью или лишь слегка выраженной болезненностью, обыкновенно легко переносимой. Однако у некоторых лиц, при слабом двигательном эффекте или даже вовсе без него, чувствительность к механическому сдавливанию локтевого нерва бывает настолько велика, что больные неохотно его переносят. Это говорит в пользу того, что возбудимость центростремительных и центробежных волокон нерва не идет параллельно одна другой. Особенно часто наблюдается повышенная восприимчивость к механическому раздражению центростремительных волокон локтевого нерва у детей.
Заслуживает внимания, что в случаях крайне резко выраженного феномена локтевого нерва я имел возможность вызывать аналогичным приемом, т. е. таким же механическим раздражением, производимым снаружи и над локтевым сочленением, двигательный эффект с лучевого нерва, сопровождающийся сокращением мышц, иннервируемых этим нервом. Точно так же можно было получить соответствующие явления со срединного нерва, сдавливая его вышеуказанным приемом снутри плечевой части руки над локтевым сочленением и даже при механическом
1 Обратный эффект ослабления двигательного эффекта до полного его прекращения также наблюдался мною, но сравнительно редко.
538
раздражении тем же приемом общего пучка нервов внутренней поверхности плечевой части руки. Наконец, в отдельных случаях я наблюдал сокращения соответствующих мышц при аналогичном механическом раздражении малоберцового нерва. Однако все же наиболее показательным вследствие легкости вызывания самого феномена должно быть признано выявление повышенной механической возбудимости локтевого нерва, что, повидимому, объясняется по крайней мере частью, более благоприятной для механического раздражения топографией этого нерва.
Интересно с этим сопоставить, что, по исследованиям Р. Я. Гол акт1, отношение порогов гальванического тетануса
/КЗТ АЗС\ к порогу
быстрых гальванических сокращений I t/gr и «от 1 >
характеризующее симпатическую иннервацию нервно-мышечного аппарата, неодинаково для различных нервов, причем это отношение является наименьшим для локтевого и малоберцового нервов и наибольшим для лицевого.
Надо заметить, что и поколачивание молоточком локтевого нерва сопровождается при повышенной его возбудимости соответствующими явлениями в отношении мышечных сокращений, но прием этот далеко не так удобен на практике, и потому в своей клинике я предпочтительно пользуюсь вышеописанным приемом механического раздражения нерва.
Давно известен симптом Хвостека, состоящий в сокращении лицевых мышц при поколачивании ствола лицевого нерва впереди уха. Спрашивается, в какой мере симптом Хвостека и мною описанный феномен являются совпадающими друг с другом. На основании целого ряда своих наблюдений могу сказать, что особого параллелизма между тем и другим явлением не существует, ибо часто имеется резко выраженный локтевой феномен, тогда как симптома Хвостека обнаружить не удается или он крайне слабо выражен. Ясно, что описываемый мною феномен является более чувствительным показателем спазмофилии, нежели симптом Хвостека, хотя при тетании имеются обычно налицо и тот и другой симптом.
Я убедился, кроме того, что очень важно отличать настоящий феномен Хвостека, состоящий в мышечных сокращениях лицевых мышц при поколачивании самого лицевого нерва впереди ушной раковины, от рефлекторного явления, которое получается при поколачивании по мышцам лица вообще, например, в области щеки. Последнее в случаях спазмофилии нередко сопровождается рефлекторными, иногда довольно распространенными, сокращениями мышц лица, захватывающими в более редких
1 Р. Я. Г о л а н т, Журнал психологии, неврологии и психиатрии. Приложения, т. 11, 1925; Р. Я. Гол ант и Я. А. Р а т н е р, Врачебная газета, 1924; Klin. Wschr., 1924, N 37.
19 В. .4. Бехтерев
289
случаях и верхние отделы лицевого нерва, тогда как самый ствол лицевого нерва ььереди уха оказывается механически не возбудимым. Дело идет здесь, очевидно, не столько о прямом сокращении ветвей лицевого нерва, сколько о рефлекторном эффекте с центростремительных ветвей тройничного нерва, который при многократном поколачивании по мышцам иногда даже начинает возрастать подобно описанной мною реакции возбуждения при тетании. С этим рефлекторным явлением, получающимся при поколачивании мышц лица, вышеописанный локтевой феномен не имеет строгого соответствия, а наоборот, даже нередко на стороне более резко выраженного феномена локтевого нерва рефлекторные явления с мышц лица на соответствующей стороне оказываются выраженными относительно слабее.
Спрашивается, в каких же случаях наблюдается феномен локтевого нерва? Прежде всего мы наблюдали его в разной степени развития у целого ряда эпилептиков, где он наблюдается с поразительной частотой и иногда представляется выраженным в крайне резкой степени. У эпилептиков мы встречаемся и с колебаниями в силе этого симптома, которые стоят в известной степени в связи с их припадками. Но и другие общие неврозы могут^ сопровождаться выявлением рассматриваемого феномена, поскольку смазмофильность является свойством их конституции. Необходимы, однако, еще дальнейшие наблюдения относительно частоты этого симптома при тех или других неврозах.
Но что заслуживает внимания —это то, что и при органических церебральных заболеваниях этот симптом может быть выражен с достаточной ясностью или даже может оказаться резко повышенным. Несколько лет тому назад я представил ряд больных с энцефалитом на конференции ленинградских врачей, у которых рассматриваемый феномен был резко выражен1. С другой стороны, я находил его резко выраженным и при джексоновской эпилепсии с односторонними судорогами и односторонним парезом на стороне последнего. В свежих случаях гемиплегии, если оно не маскируется контрактурой, мы также встречаемся иногда с повышением локтевого феномена. Эти факты вполне объяснимы, если подходить к описанному явлению с точки зрения спинномозгового рефлекса, который, естественно, должен повышаться при недеятельности и перерождении двигательного пути.
Отсюда очевидно, что повышение локтевого феномена может зависеть, с одной стороны, от гормональных условий в случаях спазмофилии и, с другой стороны, от особенной иннервации двигательного цереброспинального пути.
В заключение следует сказать, что и в других случаях (например, при эпилепсии и общих неврозах) локтевой феномен представляется далеко не редко неравномерным, и причем более резким он оказывается обычно на стороне более резко выражен-
1 В. Бехтерев, Конференция врачей по энцефалиту в 1924 г.
290
ной симпатикотонии, характеризующейся расширением зрачка на этой же стороне и другими явлениями; наоборот, вышеуказанный рефлекс с мышц лица представляется повышенным часто на противоположной стороне.
Все вышеизложенное заставляет нас притти к следующим выводам:
1. Описанный мною двигательный феномен локтевого нерва, получающийся при механическом раздражении локтевого нерва, представляет собою явление смешанного характера и обусловливается частью прямым механическим раздражением локтевого нерва, на счет чего следует поставить сгибательное движение V и IV пальцев, частью является рефлекторным актом, обусловленным передачей возбуждения с центростремительных проводников этого нерва через спинной мозг, на счет чего следует поставить сгибательное движение других пальцев руки и сокращение целого ряда мышц, иннервируемых другими нервами.
2. Феномен локтевого нерва при отсутствии органических поражений обычно является выражением общей спазмофильности и потому стоит в прямой зависимости от гормональных условий, связанных, повидимому, прежде всего с недостаточным отделением околощитовидных желез.
3. При односторонних парезах и параличах феномен локтевого нерва, где он существует вообще, представляется сильнее выраженным на соответствующей стороне, что следует отнести на счет недеятельности и перерождения центрального двигательного пути.
4. Феномен локтевого нерва является для огромного ряда случаев более чувствительным показателем спазмофильности, нежели симптом Хвостека на лицевом нерве и рефлексы с лицевых мышц.
5. В резких случаях спазмофильности феномен локтевого нерва может нарастать при многократном механическом раздражении нервного ствола.
6. Феномен локтевого нерва в случае его неравномерности всегда более выражен на стороне более резко выраженной симпатикотонии.
7. От двигательного феномена локтевого нерва следует отличать «чувствительный» локтевой феномен, который служит выражением повышенной возбудимости центростремительных волокон локтевого нерва.
8. При чрезмерно повышенной спазмофильности аналогичные явления двигательного и чувствительного характера могут быть обнаружены при механическом раздражении и других нервов, например, лучевого, срединного, малоберцового и т. п.
19*
ТАХИФЕМИЯ, ТАХИБАЗИЯ И ПАРОКСИЗМАЛЬНОЕ
ТОПТАНИЕ НА МЕСТЕ КАК ПОСТЭНЦЕ-
ФАЛИТИЧЕСКИЙ СИМПТОМ
В научной литературе мало выясненным представляется пост-энцефалитический симптом, характеризующийся прогрессирующим или временным ускорением темпа движений. Сколько я мог проследить за все более и более растущей литературой энцефа-литических и постэнцефалитических расстройств, до сих пор были опубликованы лишь одиночные наблюдения с обращением Бнимания на вышеуказанное расстройство движений. Один такой случай с подробным выяснением природы этого расстройства был опубликован И. Герстманом и П. Шильдером1, где водном случае у постэнцефалитической пациентки наблюдался этот симптом в отношении речи: больная начинает говорить нормальным темпом, затем происходит очень быстрое ускорение речи, которое становится все более непреодолимым и тягостным, причем ни артикуляция, ни содержание речи при этом не нарушаются. Поразительно, что пациентка в отношении такой области, как еда и жевание, представлялась акинетичной. Заслуживает упоминания, что при особом напряжении внимания она могла на короткое время подавлять этот симптом. Вместе с тем общее поведение пациентки с характером своеобразной привязчивости юношеских энцефалитиков, отличающихся чрезмерной подвижностью, распространялось и на речь.
Из французских авторов Клод в Парижском неврологическом обществе заявляет о расстройстве речи под названием прогрессивной tachyphemie paroxystique в числе энцефалитических симптомов. Дело идет в этом случае об ускорении речи с повторением слов и фрагментов фраз и в быстром его нарастании до полной их неразборчивости. Симптом этот наблюдался как при передаче прошлых происшествий, так и при счете.
Равным образом и мне приходилось наблюдать подобное же явление при послеэнцефалитическом заболевании. В моем случае при явлениях амимии и паркинсонизма тахифемия проявлялась каждый раз, как больной начинал говорить, отвечая на вопросы.
1 I. Gerstmann u. P. Schilder, Zschr. f. d. ges. Neurol, u. Psych., Bd. 81, Hft. 4 — 5, S. 570 — 582.
292
Начав свою речь обыкновенным темпом, больной затем начинал произносить ответ с каждым словом все скорее и скорее и, наконец, произносил начальный слог последнего слова, повторяя его многократно, наподобие постепенно затихавшей барабанной дроби, чем обыкновенно и заканчивался его ответ.
Здесь дело шло, очевидно, о прогрессивном ускорении темпа речи, воздержаться от которого больной не был в состоянии никаким образом. Совершенно аналогичное явление я наблюдал и в другом, подобном вышеуказанному случае.
И в том, и в другом случае обращало на себя внимание то обстоятельство, что сами больные вовсе не были словоохотливыми, наоборот, больные были обычно молчаливы, их уста были скованными так же, как и другие части тела, но, начав говорить, они как бы выпаливали словами, преодолевая скованность речевых мышц, после чего каждое произносимое слово является как бы стимулом для ускорения следующих слов в той же фразе.
Дело идет, таким образом, о больных, у которых, наряду с явлениями общей скованности и напряженности мышц (паркинсонизм) и одновременной акинезии, наступают расстройства речи с характером прогрессирующего ее ускорения до той степени, что произношение целых слов становится уже невозможным н, в конце концов, начинает повторяться только один слог при постоянном затихании голоса. При этом самое произношение отличается все время своеобразной однотонностью голоса. Ясно, что здесь при однотонности речи имеется акинезия голоса и речевых движений вообще и вместе с тем ускорение темпа речевых движений до автоматического повторения скороговоркой одного и того же слога при постепенном замирании голоса. Однотонность речи следует объяснить скованностью или статическим напряжением мышечного аппарата, что может стоять в связи с устранением тормозящих влияний, осуществляемых церебральными связями мозжечка. Ускорение темпа речи может быть поставлено также в связь с ослаблением тормозных процессов, играющих роль в каждом вообще движении, при котором сокращение одной группы мышц сменяется сокращением другой группы.
Нужно затем иметь в виду, что начальные речевые движения, подкрепляемые, как и все вообще движения, произвольной иннервацией, отличаются в смысле известного усилия достаточными двигательными импульсами, тогда как дальнейшее произношение слов, осуществляемое более автоматично, постепенно все более и более слабеет вследствие имеющейся акинезии движений вообще.
Эта акинезия, или ослабление нервной иннервации, ввиду отсутствия признаков участия в поражении пирамидных путей, очевидно, должна быть поставлена в этих случаях в связь с парс-тическим состоянием внепирамидных двигательных проводников, проходящих через стриальные образования.
293
Последние, как я высказался уже много лет тому назад в своих «Проводящих путях» (СПБ, изд. Риккера, т. 2), представляют внутреннюю извилину мозга, протягивающуюся от переднего дырчатого пространства в нижний рог до миндалевидного ядра височной доли, которая, как мы можем установить, имеет, подобно другим корковым областям, парную систему центростремительных и центробежных проводников. Первая идет от полукружных каналов, поднимаясь с преддверным нервом и его дальнейшими продолжениями в мозжечке и в его передней ножке, и от периферических нервно-мышечных приборов, поднимающихся по спинному мозгу через мозжечок и его переднюю ножку, красное ядро до стриопаллидальных образований и лобной доли мозга; центробежная же система идет через лобно-мостовые системы и описанный мною центральный пучок покрышки.
На основании своих наблюдений я убедился, что явления, аналогичные тому, что мы наблюдали в речи больных, могут быть наблюдаемы при постэнцефалитическом паркинсонизме и в отношении ходьбы. Такого рода больные, начав ходьбу с нормальной скоростью, все более и более ускоряют темп смены своих шагов и, кажется, вот-вот могут упасть от непрерывного ускорения передвижения, но они успевают обыкновенно достичь стены или другого какого-либо препятствия, в которое они и упираются с известной стремительностью. Есть основание полагать, что это болезненное явление, которое можно бы назвать как трафтом, стоит в прочной связи с антеропульсией, ибо во всех такого рода случаях оба явления обнаруживались большей частью совместно.
Вообще в походке паркинсоников, как и в данном случае, обращает, между прочим, особое внимание тот факт, что эти больные, начав ходить медленно, все более и более ускоряют свое движение. Это можно было бы объяснить тем, что ригидность, развивающаяся в статическом положении, лишь постепенно уступает динамизму мышц. Но в данном симптоме, несомненно, играет роль и ослабление тормозных влияний, ибо, как известно, в этих случаях больные сами не могут остановить своего движения. В этих случаях механизм ходьбы, начавший работать, не может быть сдержан самим больным, несмотря на то, что он вводится в действие не без некоторого активного усилия.
Если, однако, по отношению к передвижению может представляться еще для кого-либо вопросом: не играет ли здесь роль перемещение в направлении кпереди центра тяжести во время ходьбы, то аналогичное явление, наблюдаемое в таких автоматических движениях, как речь, не может оставлять никакого сомнения в отношении недостаточности тормозов по отношению к развернувшемуся автоматизму.
Что в непрерывном ускорении ходьбы играет роль самый механизм ходьбы, а не нарушение равновесия, сопровождающееся
294
произвольными усилиями с целью подпереть перемещающийся вперед центр тяжести тела, доказывает, между прочим, тот факт, что такого рода больные лучше ходят не по ровной поверхности пола, а при том случае, если больным приходится перешагивать через препятствия. Это своеобразное явление обратило мое внимание лишь в самое последнее время, а потому я не могу сказать, насколько оно отличается постоянством в подобного рода случаях. Но во всяком случае оно может быть крайне резко выраженным. Вот, например, больной, около 45 лет, с резко выраженным paralysis agitans с явлениями паркинсонизма с отсутствием указаний на энцефалитическое происхождение. Лицо его маскообразно, речь монотонная, верхние конечности слегка полусогнуты в локтевых сгибах, в руках резко выраженное дрожание, отсутствие патологических рефлексов в нижних конечностях, больной представляет явления тонической астазии, но тем не менее он поднялся ко мне во второй этаж, пройдя свободно два марша лестницы. Когда я предложил ему пройтись по комнате, он начал быстро ускорять свою ходьбу, пока не уперся почти с разбегу в противоположную стену. Но он сам обратил мое внимание на то, что он гораздо лучше ходит при условии, если ему приходится переходить через препятствия.
Тотчас же было приступлено к проверке этого заявления. Для этой цели были расставлены по направлению его ходьбы четыре стула в боковом положении, и оказалось, что он отлично прошел правильно то же расстояние, перешагивая через каждый стул, ставя свои ноги между ножками стульев и выводя их оттуда, причем в этом случае не обнаруживалось и постоянного ускорения ходьбы, которое всегда наблюдается у больного при обыкновенном беспрепятственном передвижении. Облегчение ходьбы при перешагивании через препятствия в данном заболевании и, очевидно, объясняется тем, что встречающиеся на пути препятствия вынуждают в гораздо большей мере участвовать в передвижении корковые импульсы, которые, будучи осуществляемы с помощью пирамидной системы, путем тормозящих влияний на автоматический механизм ходьбы улучшают его работу, замедляя темп движения.
Но есть и еще своеобразный тип расстройства в передвижении больных с послеэнцефалитическим процессом, который мне встретился пока в небольшом числе случаев, но в чрезвычайно эксквизитной форме. В некоторых случаях у больных, перенесших энцефалит, наблюдается симптом непреодолимого многократного переступания с ноги на ногу во время ходьбы, пере-ступания, напоминающего шаг на месте и наблюдаемого время от времени при передвижении, особенно же с начала ходьбы, а также перед порогом, который больные должны перешагнуть, затем при поворотах. Таким образом, у некоторых больных имеется своеобразный симптом, который характеризуется тем, что больной, начав ходить обычным по размеру шагом, начинает
295
совершенно неожиданно семенить, быстро проделывая некоторое время небольшие шажки почти на одном месте или в пределах небольшого пространства, после чего больные снова начинают ходить обыкновенным шагом.
Так как такое явление в походке чаще всего обнаруживается в случаях необходимости делать поворот во время ходьбы и в случае необходимости пройти через двери, перешагнуть через порог и т. п., то, повидимому, это изменение походки является результатом каких-то импульсов, исходящих из коры мозговых полушарий; этот симптом пароксизматического быстрого переступания с ноги на ногу наблюдался мною уже в нескольких случаях и всегда со стереотипным постоянством. Поразительно, что симптом этот, повидимому, до сих пор не отмеченный в научной литературе, я встретил в одном случае, где выступали с особенною резкостью астенические явления, явившиеся следствием бывшего ранее энцефалитического заболевания.
О СЛОЖНЫХ РЕФЛЕКТОРНЫХ ЯВЛЕНИЯХ В НИЖНИХ
КОНЕЧНОСТЯХ ПРИ ПОРАЖЕНИЯХ ЦЕНТРАЛЬНОГО
ДВИГАТЕЛЬНОГО НЕВРОНА
(Рефлексы спинномозгового автоматизма)
За последние полтора-два десятка лет стали заниматься особенно много рефлексами спинномозгового автоматизма (reflexes d'automatisme medulla ire) и в том числе так называемыми защитными рефлексами (reflexes dits«de defense»). Этим явлениям были посвящены работы П. Мари и Ш. Фуа1, Андре Томаса2, И. Бабинского, Венсана и Жарконского3, В. фон Веркома4, И. Маринеско и Д. Нуака5, И. Бабинского и Жарконского6, Андре Строля7 и многих других.
Ввиду этого интереса к рефлекторным явлениям, имеющим особую практическую важность, считаю необходимым указать,, что в русских невропатологических клиниках вышеуказанные рефлекторные явления уже давно вошли в методику исследования нервнобольных, благодаря исследованиям, которые были сделаны мной значительно раньше, чем появились в литературе вышеуказанные работы французских авторов.
Так, еще в 1900 г. в русской литературе я опубликовал сообщение под заглавием «о сложных сухожильно-мышечных реф-
1 P. Marie et Ch. Foix, Les reflexes d'automatisme medul-laire et le phenomene des raccourcisseurs etc., Revue neurol., mai 1912; Онн же, Reflexes d'automatisme medullaire et reflexes dits «de defense» etc., Sem. med. du 22 oct 1913.
2 A η d r 6 Thomas, La paraplegie spasmatique avec contracture variable etc., La clinique, 20 et 27 juin, 1913.
3Babinski, Vincent etJarkonski, Les reflexes cutanes de defense dans la maladie de Friedreich, Societe de neurol de Paris, Seance du 7 mars 1912.
* W. von Woerkom, Sur la signification physiologique des reflexes cutanes des membres inferieurs etc., Revue neurol., 15 sept. 1912.
5 I. Marinesco et D. Noica, Sur les reactions des membres inferieurs etc., Revue neurol., 30 avril 1913.
•I. Babinski et Jarkonski, Sur la possibilite de determiner la hauter de la lesion dans les paraplegies etc., Societe de neurol. de Paris, 12 mai 1910; Revue neurol., 30 mai 1910.
'Andre Strohl, Les reflexes d'automatisme medullaire, Paris, 1913.
297
лексах нижних конечностей при миэлитах1. В этом сообщении я описываю при повышенной рефлекторной возбудимости нижних конечностей разгибательный рефлекс ноги, при котором «одновременно и совместно сокращаются как разгибатели бедра, так и икроножные мышцы, как будто бы это была одна группа мышц, выполняющая определенное движение». Этот рефлекс «легче и удобнее всего вызывается быстрым пассивным сгибанием ноги в колене, при котором разгибатель бедра подвергаются пассивному растяжению». Но «иногда этот рефлекс удается вызывать и тыльным сгиба1 ием стоп, приводящим в пассивное растяжение икроножные мышцы». Очевидно, что это пассивное и притом всегда лишь быстрое растяжение разгибателей и служит раздражителем, возбуждающим этот рефлекс, между тем как медленно производимые пассивные сгибания бедра при вытянутой стопе удаются свободно, не вызывая разгибательного рефлекса.
Другим, так же часто наблюдаемым рефлексом, хотя и менее постоянным, является «сгибательный рефлекс ноги»; «этот рефлекс выражается одновременным сокращением сгибателей бедра и сгибателей голени (tibialis anticus, peroneus), вследствие чего нога сгибается в коленном сочленении, стопа же и пальцы, особенно большой, производят тыльное сгибание». «Движение это вызывается легче всего подошвенным сгибанием стопы, но оно также легко может быть вызвано простым сжатием бедренных и икроножных мышц между пальцами руки, а иногда даже простым смещением коленной чашки вытянутой ноги, вызывающим растяжение разгибателей бедра, которое, очевидно, рефлекторным путем передается на сгибатели конечностей. И здесь «сгибатели бедра и стопы действуют совместно при вызывании выше указанного сгибательного рефлекса ноги».
«В некоторых случаях даже тыльное сгибание стопы вызывает то же движение, благодаря, очевидно, тому, что происходящее при этом растяжение икроножных мышц рефлекторно вызывает сокращение антагонистов, главным образом tibialis anticus и peronei. В последнем случае вслед за произведенным сгибанием стопы вместе с возвращением стопы в прежнее положение обнаруживается тотчас же разгибательное движение пальцев ноги, особенно большого, причем ясно напрягается и сухожилие последнего». Это рефлекторное движение в сущности представляет собою так называемое«парадоксальное сокращение».
«Наконец, третьим сложным рефлексом является ,,приводящий рефлекс ноги", который получается в том случае, когда ноги больного при полусогнутом положении колена быстро и пассивно разгибаются. Этим самым приводятся в пассивное растяжение аддукторы бедра, что приводит тотчас же к рефлекторному их сокращению, благодаря чему обе нижние конечности приво-
1 Обозрение психиатрии, 1900, № 9.
298
дятся. Рефлекс этот большею частью, впрочем, представляется далеко не столь резко выраженным, как разгибательный и сгиба-тельный рефлекс нижних конечностей, и встречается в общем сравнительно редко».
Достойно внимания, что вышеуказанные рефлексы, особенно первые два, при известных условиях могут быть вызываемы и кожными раздражениями, например, раздражением с помощью губки, смоченной холодной водой, или щипком кожи нижних конечностей, а равно и другими кожными раздражениями.
В то время как писались эти строки, были известны только некоторые данные Штрюмпеля, сообщенные им в докладе «Кожные и сухожильные рефлексы при заболеваниях нервной системы» (собрание юго-западных неврологов и психиатров в Баден-Бадене 3 и 4 июня 1899 г.) и затем phenomene des orteils, Бабинского, рефлекс Шеффера и некоторые другие. Обсуждая разногласие между обоими этими авторами относительно природы рефлекса Шеффера1, в той же работе я говорю: «нетрудно видеть, что рефлекс антагонистов, описанный Шеффером и отождествляемый Бабинским с описанным им phenomene des orteils в сущности представляют собою тот же тип рефлекторного движения, который мы наблюдаем при нашем разгибательном и сгибательном рефлексах нижних конечностей». Но эти рефлексы вызываются, как мы видим, в одних случаях мышечными раздражениями (сдавливанием их и растяжением), в других случаях кожными раздражениями.
Из этой работы ясно, что еще в 1900 г. мной был описан как Ie phenomene des allongeurs, так и phenomene des raccourcis-seurs, причем мной указан был и прием сильного сгибания пальцев стопы, вызывающий появление этого рефлекса, а также иные приемы, приводящие к появлению тех же рефлексов.
Позднее к этому способу вызывания сгибательного рефлекса ноги, ныне обычно обозначаемого французскими авторами защитным рефлексом (reflexe de defense), я возвращаюсь в других работах, помещенных и в русской в 1905 г.2 и немецкой печати в 1906 г.3.
Я привожу здесь текстуально то, что было сказано мной по поводу этого рефлекса в только что указанной работе, опубликованной в «Neurologisches Zentralblatt» в 1906 г.
«В числе признаков органического паралича центрального неврона нужно отметить одно явление, которое наблюдается
1 Soc. neurol. de Paris, 11 janv. 1900.
2 В. Бехтерев, О рефлекторных явлениях при органических параличах центрального неврона и о новом тыльно-стопном сгибательном рефлексе ноги, Обозрение психиатрии, 1905, стр. 771.
3 В. Бехтерев, Ueber eine eigentumliche Reflexerscheinung bei Plantarflexion des Fusses und Zehen in Fallen von Affektion des zentra-len motorischen Neurons, Neurol. Zbl., 1906, N 7, и Обозрение психиатрии, 1906.
299
иногда при патологически повышенной рефлекторной возбудимости. Это явление заключается в следующем: если взять в руки стопу больного и произвести сильное подошвенное сгибание как ее самой, так и пальцев, то вслед за таким подошвенным сгибанием стопы и пальцев происходит тотчас же тыльное сгибание стопы и пальцев, а в более резко выраженных случаях и при более сильном подошвенном сгибании стопы вместе с последовательным тыльным сгибанием стопы и пальцев происходит даже сгибание ноги в коленном и бедренном суставах».
Здесь дело идет, очевидно, о своеобразном рефлекторном явлении, развивающемся вследствие растяжения сухожилий тыльных сгибателей стопы и разгибателей пальцев, и приводящем к сокращению соответствующих мышц, вследствие чего и происходит последовательное тыльное сгибание стопы и пальцев. При этом в случае более сильного подошвенного сгибания стопы и пальцев рефлекс распространяется не только на те мышцы сухожилия, которые подвергаются растяжению, но и на сгибателей колена и бедра.
Рассматриваемый феномен вызывается вообще без труда там, где он имеется. До сих пор он наблюдался мною при травматических поражениях спинного мозга, при спинномозговом сифилисе, при миэлитах с резким повышением рефлексов; равным образом он наблюдается иногда и при церебральном поражении двигательного неврона.
Служа выражением общего повышения рефлекторной возбудимости, рассматриваемый феномен должен быть принимаем во внимание при диагнозе поражений, захватывающих те или другие части центрального двигательного неврона наряду с другими явлениями, как клонус стопы, рефлекс Бабинского, феномен Оппенгейма, описанное мною и затем К. Менделем подошвенное сгибание пальцев при постукивании молоточком по тылу стопы, описанное мною нарастание рефлекторной возбудимости при вызывании костного рефлекса частым постукиванием молоточком и т. п.
«Уступая по частоте появления клонусу стопы и рефлексу Бабинского, вышеописанный феномен имеет особенную ценность как указатель поражения центрального неврона там, где вследствие контрактуры и ригидности мышц нижних конечностей не удается вызвать в них ни клонуса стопы, ни даже рефлекса Бабинского, тогда как вышеуказанный феномен может обнаруживаться с ясностью».
Таким образом, из предшествующих работ, из которых первая относится к 1900 г., а другие к 1905—1906 гг., можно вполне убедиться, что общий сгибательпый рефлекс был мною уже в самом начале текущего столетия не только описан, но и оценен по достоинству в смысле его диагностического значения как указателя на органическое поражение центрального двигательного неврона.
300
Поэтому когда в Казанском обществе невропатологов и психиатров было сделано указание об этом рефлексе на основании данных Фуа доктором Фаворским, что сочлен этого же общества, бывший ассистентом клиники нервных болезней Казанского университета, доктор И. А. Веселитский1 сделал в русской печати в особой статье под заглавием «Рефлекс будто бы П. Мари и Фуа» сообщение следующего содержания (привожу, конечно, содержание статьи в кратком извлечении).
«В заседании 7 июля 1910 г. Общества неврологии в Париже (Societe de neurologie de Paris) П. Мари и Фуа демонстрировали (вернее, демонстрировал собственно Фуа) новое наблюдавшееся ими явление, рефлекс оттягивания нижних конечностей при некоторых поражениях центрального неврона двигательного пути. Сообщая об этом обществу, авторы, очевидно, находились в приятном заблуждении, что этот рефлекс наблюдался ими впервые, так как иначе они не могли бы умолчать о прежних наблюдениях, сюда относящихся. Феномен этот так представлен авторами, судя по отчетам заседания. Они сообщают, что при более или менее форсированной флексии пальцев ноги с трансвер-зальной прессией стопы нормально не вызывается никакого движения; в случаях же поражения пирамидного пути при этом наблюдается: 1) флексия бедра к тазу, 2) флексия голени к бедру и 3) флексия стопы к голени с приведением ее и ротацией внутрь.
Доктор Фаворский, имевший удовольствие во время своей заграничной командировки видеть этот новый рефлекс, так сказать, в собственном исполнении П. Мари, под свежим впечатлением и несомненным обаянием таланта этого ученого, поспешил поделиться этой новинкой в первом же заседании общества невропатологов и психиатров по возвращении своем из-за границы.
Слушая это сообщение, я никак не мог отделаться от смутных воспоминаний, вызванных им, что как будто что-то совершенно подобное когда-то и где-то мне уже встречалось; что тут дело идет о чем-то уже известном и знакомом. Несмотря на все усилия восстановить с полной ясностью эти смутные воспоминания, это мне не удалось именно тогда, когда нужно — явление, обычное у людей моего возраста. Конечно, об этих мучащих меня сомнениях я говорить в обществе не мог и сделал некоторые другие замечания по поводу сообщения, о которых речь будет ниже. На другой же день после доклада мне пришлось с очевидностью убедиться, что мои смутные воспоминания меня не обманули. Я довольно скоро нашел коротенькое сообщение о том же предмете нашего почтенного академика В. М. Бехтерева, помещенное в распространенном немецком журнале (см. ,.Обозрение Психиатрии и пр.", 1905, стр. 571 и „Neurologi-sches Zentralblatt" за 1906 г.,№ 7, стр. 290), где описан как раз
1 Д-р Веселитский, Обозрение психиатрии, 1910, № 12.
301
тот же рефлекс, который через четыре года мне пришлось встретить в нашем обществе под другой этикеткой в сообщении доктора Фаворского, в качестве совершенно свежей новости. Для убедительности позволю себе сделать здесь несколько выдержек из упомянутой статьи В. М. Бехтерева. Уже самое заглавие „Ueber eine eigentumliche Reflexerscheinung bei Plantarflexion des Fusses und Zehen in Fallen von Affektion des zentralen moto-rischen Neurons" доказывает, что дело здесь идет как раз о том, что в этом году наблюдали впервые П. Мари и Фуа.
Дальнейшее убеждает в этом еще более и представляет только незначительную разницу в приемах, способах, но с одной и той же сутью дела. Между симптомами органического паралича центрального неврона, говорит В. М. Бехтерев, достойно внимания одно явление, которое наблюдается иногда при патологически повышенной возбудимости рефлексов... если обхватить стопу больного и приводить ее, как и пальцы, в форсированное подошвенное сгибание, то за этим следует тотчас дорзальное сгибание стопы и пальцев, а при более форсированном подошвенном сгибании — также сгибательное движение в колене и бедре. Автор дает краткие объяснения сути этого феномена, где, с одной стороны, приходят в сокращение мускулы, сухожилия которых подвергаются растяжению и дают дорзальное сгибание стопы и пальцев, с другой —последовательно вызывается сокращение мускулов вышележащих, сухожилия коих не подвергаются растяжению. Затем перечисляет те случаи, где ему приходится наблюдать этот феномен: травматическое поражение спинного мозга, спинальный сифилис, миэлиты и иногда церебральные заболевания. В заключение он выражает надежду, что наблюдаемый симптом как выражение общего повышения рефлексов может быть полезен при диагнозе поражений центрального двигательного неврона и может быть поставлен наряду с другими симптомами: клонус стопы Бабинского, Оппенгейма, пальцевой рефлекс его же —Бехтерева, к которому как-то удалось пристроиться и Курту Менделю. Этот феномен, по его мнению, хотя и стоит позади упомянутых других, в некоторых случаях, соединенных с контрактурой и ригидностью, вызывается легче.
Из этого сопоставления сообщений академика В. М. Бехтерева (четыре года тому назад) и сообщения П. Мари и Фуа в этом году не подлежит ни малейшему сомнению, что дело тут идет об одном и том же явлении и что будто бы только что открытый П. Мари и Фуа феномен на самом деле уже был раз открыт и притом четыре года тому назад В. М. Бехтеревым1. Отсюда явствует, что этикетку, под которой он явился в обществе невропатологов и психиатров в Казани в изложении доктора Фаворского, надлежит снять и заменить ее более соответствующей и
В действительности, как видно из вышесказанного, более 10 лет назад, т. е. ранее описания Фуа и П. Мари.
302
справедливой, т. е. именовать, если это нужно, феноменом В. М. Бехтерева. На долю парижских наблюдателей в этом деле досталось только подтверждение (хотя и самостоятельно) верности наблюдения В. М. Бехтерева и некоторые несущественные подробности с некоторой модификацией приемов, на мой взгляд, менее удачных. Модификация эта заключается в применении вместо подошвенной флексии стопы трансверзальной прессии ее, что, по свидетельству самих Мари и Фуа, представляется менее чувствительным приемом и не всегда ведет к цели. Я думаю, что этим актом справедливости, т. е. восстановлением несомненного приоритета В. М. Бехтерев относительно описываемого феномена, мы, конечно, нисколько не умаляем высокое положение,, бесспорно занимаемое такой крупной величиной в невропатологии, как П. Мари. Напротив, критическое отношение, а не рабская слепая вера во все, что исходит от таких деятелей, только может повысить интерес к их исследованиям, а не умалять их». Лично я, по получении реферата о прочитанном Фуа сообщении в Парижском неврологическом обществе, тотчас же выслал П. Мари и Фуа отдельный оттиск моей работы из «Neurologi-sche's 2епгга1ЫаИ:»для их ознакомления с указанием на страницы руководства Левандовского, в котором упомянутый рефлекс был также описан на основании моих исследований. К сожалению, ни письменного отклика на мою посылку, ни ссылок в позднейших работах (Фуа и П. Мари) на мои исследования по указанному предмету я не встретил.
Руководясь этим, в 1911 г. специально для ознакомления русских врачей-неврологов с описанными мною рефлексами и в том числе со сгибательным рефлексом ноги, вызываемым с помощью подошвенного сгибания стопы и ее пальцев, было сделано мною в обществе врачей клиники душевных и нервных болезней1 сообщение с демонстрацией нескольких больных, у которых упомянутый симптом представлялся резко выраженным и давал определенные диагностические указания о характере центрального поражения. Одновременно были демонстрированы и больные с защитными рефлексами при уколах в тыл и другие части стопы, а также рефлекс подобного же рода, получающийся при многократном постукивании врачебным молоточкам по тылу стопы.
Вслед за этим мной было опубликовано в том же году исследование под заглавием: «О рефлекторных явлениях при органических параличах центрального неврона и о новом тыльно-стоп-ном рефлексе ноги», которое было помещено мною и в русской2, и в немецкой печати3.
В этой работе, кроме дальнейших указаний насчет того же рефлекса, вызываемого насильственным сгибанием пальцев,
1 См. отчеты этих собраний за вышеуказанный год.
2 В. Бехтерев, Обозрение психиатрии, 1911, № 7. 8 W. В е с h t e r e w, Neurol. Zbl., 1911.
303
я отмечаю уже упомянутое при демонстрации особое явление рефлекторного характера, выражающееся тем же сгибательным рефлексом ноги с отгибанием к тылу стопы и большого пальца, но вызываемом многократным постукиванием врачебного молотка по тылу стопы. В этом случае уже «после нескольких ударов стопа обнаруживает тыльное сгибание, пальцы, особенно большой, разгибаются и происходит сгибательное движение всей йоги в тазобедренном, коленном и голено-стопном суставе».
В упомянутой работе я указываю, что и ранее описанный мною при центральных параличах сгибательный рефлекс ноги, получающийся при многократном постукивании врачебным молоточком по передней поверхности большеберцовой кости, представляет собою аналогичное с вышеуказанным рефлексом явление.
Кроме того, я указываю, что описываемый сгибательный защитный рефлекс ноги с отгибанием стопы и большого пальца к тылу вызывается не только вышеуказанными приемами, но еще и «уколами в области тыла стопы» —явление, которое было демонстрировано мною не только в указанном сообщении в обществе клинических врачей психиатров и неврологов, но и много ранее того па своих клинических лекциях.
При этом поразительным является то обстоятельство, что в последнем случае «рефлекс не носит характера движения, отстраняющего орудия, а напротив того, как бы направляющего стопу на колющее орудие».
Надо, впрочем, заметить, что уколы и в другие части стопы, например, боковые части и пр., не говоря об уколах в подошву, способны вызывать появление того же самого сгибательного рефлекса, но с отгибанием к тылу стопы и большого пальца, который вызывается и штриховым раздражением подошвы стопы в виде известного феномена Бабинского.
В остальном работа касается клинического значения вышеуказанного рефлекса, который, как я говорю в упомянутой работе, «принадлежит, вообще говоря, к числу более или менее постоянных признаков параличей центрального неврона, обнаруживаясь как при миэлитах и других поражениях спинного мозга выше поясничного утолщения, так и при поражениях головного мозга (размягчение, кровоизлияние и пр.), прерывающих двигательные пути».
При этом я отметил тот факт, что вышеуказанный рефлекс стопы может быть получен при поражениях пирамидного пути иногда даже и в тех случаях, когда не имеется налицо ни ярко выраженного рефлекса Бабинского, ни описанного мною сгибательного пальцевого рефлекса, ни даже клонуса стопы. Этим определяется его диагностическое значение в подходящих случаях.
Ввиду всего приведенного в России вызывает только удивление, что описанные мною рефлексы во французской литера-
304
.туре обозначают рефлексами Фуа и П. Мари, несмотря на то, что эти авторы обратили внимание на этот рефлекс много лет спустя после меня. По примеру французов это заблуждение переносится и в другие страны, что и вынуждает меня еще раз осветить этот вопрос в печати. Дополнительно к сделанным мною ранее сообщениям нахожу нужным добавить, что в отношении внешнего выявления рефлекса от подошвенного сгибания стопы и пальцев мои данные стоят в согласии с результатами исследований Гольдфлама1, ибо в хорошо выраженных случаях наступает с постоянством не только сгибание стопы, колена и бедра, но и большого пальца. Этот рефлекс в моих случаях наблюдался со всеми признаками в случаях пирамидных поражений как церебрального характера (гемиплегии), так и при спинномозговых поражениях над поясничным утолщением, таких, как миэлит, рассеянный склероз и др. В случаях сдавления спинного мозга (опухоль, смещение позвонков) рассматриваемый рефлекс выражен был в моих случаях чрезвычайно ярко. В случаях коматозных состояний его также можно наблюдать, но в очень слабой степени. В нормальном состоянии рефлекс этот не наблюдается вовсе.
Вообще все мои наблюдения приводят к тому, что здесь мы имеем дело с рефлекторным явлением, связанным с дегенерацией пирамидных путей, ибо ни в одном случае с поражением вне пирамидных путей его наблюдать не удалось. В этом смысле рассматриваемый рефлекс должен быть поставлен в семью стопных патологических рефлексов, наблюдаемых в случаях поражений пирамидного пути.
Наряду с вышеуказанным рефлексом, заслуживает не меньшего внимания отмеченное выше рефлекторное явление, которое наблюдается в случаях поражения пирамидных путей и которое вызывается уколами в ту или другую часть стопы. Вполне понятно, что если мы произведем укол в подошву стопы, то произойдет соответствующий защитный рефлекс, характеризующийся отдергиванием стопы, сгибанием колена и бедра с заметным отведением последнего. Но в случаях поражения пирамидного пути мы получаем при уколе подошвы стопы вместе с резко выраженным тыльным сгибанием стопы еще и тыльное же отгибание большого пальца. Интереснее всего, что и при уколе в тыл стопы последняя производит резкое тыльное сгибание вместе с отгибанием к тылу большого пальца (а иногда и остальных) сгибанием колена и бедра с легким отведением последнего, вследствие чего, вопреки природе защитных реакций, обычно отстраняющих орган от колющего орудия, в данном случае стопа как раз отгибается в сторону колющего орудия. Дело идет здесь, таким образом, о явлении, явно не соответствующем природе
1 G о 1 d f 1 а ш, Ein Beitrag zur Semiologie d. Verkiirzungsreflexes, Dtsch. Zschr. f. Nervenheilk., 1923, Bd. 80, Hft. 3/4, S. 238 — 269.
20 В. М. Бехтерев
305
рефлексов, вызываемых кожными болевыми раздражениями, которые в нормальном состоянии всегда имеют защитный характер и приводят к устранению соответствующего органа от колющего, как и всякого другого разрушающего раздражения. Очевидно, что патологически извращенный характер рефлекса в этом случае обусловливается тем, что проведение нервного импульса здесь происходит по установившемуся пути наименьшего сопротивления.
Что касается защитного рефлекса, получающегося при параличах центрального двигательного неврона при многократном постукивании по тылу стопы врачебным молоточком, то дело идет здесь о рефлексе, обусловливаемом суммарным раздражением костей тыла стопы, которое, постепенно нарастая, достигнет той степени, когда разрешение рефлекса становится неизбежным» Но и здесь рефлекс, как и в предыдущих случаях, выявляется в определенной форме в виде тыльного сгибания пальцев, особенно большого, тыльного сгибания стопы, сгибания колена и бедра с отведением последнего, благодаря обособлению рефлекторного участка спинного мозга от тормозящих двигательных областей мозговой коры. Что дело идет здесь о суммировании раздражения, очевидно из того, что те же удары молоточком, произведенные в одиночку, ни производимые, хотя и многократно, но с сравнительно редкими ударами, неспособны вызывать подобного же рефлекса.
На появление костных рефлексов от суммарных раздражений при параличах центрального двигательного неврона я обратил внимание еще более трех с половиной десятков лет тому назад и в свое время демонстрировал и описал эти рефлексы, вызывая их многократными последовательными ударами молоточка по передней поверхности большеберцовой кости (tibia). В этих случаях рефлекс, состоящий в откидывании голени кпереди, может вовсе не получаться при однократном постукивании молоточком по передней поверхности большеберцовой кости, но если мы будем производить последовательные многократные удары молоточком по одному и тому же месту tibiae, то при параличе центрального неврона, например, в случае центральной гемипле-гии или при миэлитическом поражении спинного мозга, мы получим через то или другое число ударов, в одних случаях через 4—6, в других, менее выраженных случаях через 16—18, отдергивание голени кпереди, между тем в нормальном состоянии, как и в случае гемиплегии, на непораженной стороне таких рефлексов от суммарных раздражений мной никогда не наблюдалось.
Как я показал в свое время, рефлекс с передней поверхности tibiae может быть вызываем, не отличается постоянством в нормальном состоянии, ибо в одних случаях он может быть вызываем одиночным ударом врачебного молотка, в других случаях он может отсутствовать. Но при церебральных гемиплегиях может
306
быть появление костного рефлекса в первом случае и повышение его во втором случае. В этих случаях мы можем наблюдать, что повторные и последовательно производимые удары молоточком по tibiae вызывают через то или другое число ударов молоточка усиленный рефлекс в виде более значительного по силе отдергивания голени кпереди.
Аналогичное явление при повторном постукивании молоточком по коленному сухожилию можно наблюдать при церебральных гемиплегиях на стороне поражения и при миэлитах также в отношении коленного рефлекса. В этих случаях, повторяя многократно удары молоточком по коленному сухожилию, мы получаем через то или другое число ударов рефлекс большей силы по сравнению с обычно вызываемым коленным рефлексом, что характеризуется большим отскакиванием кпереди голени. Последняя в таких случаях как бы подпрыгивает кпереди. В более редких случаях многократное механическое раздражение коленного сухожилия приводит к тому, что голень совершенно расправляется вследствие тонического напряжения прямой мышцы бедра.
Совершенно аналогичные явления мы можем наблюдать иногда и при гемиплегиях и при многократном ударе молоточком по локтевому сухожилию. И в этом случае предплечье при многократно производимых ударах по сухожилию biceps время от времени подпрыгивает с большей силой по сравнению с тем, как это наблюдается обычно.
В ряде случаев я наблюдал, что при таком способе исследования рефлекс сопровождался тоническим напряжением сокращающихся мышц, благодаря чему многократными ударами молоточком предплечье удерживалось в горизонтальном положении, принимая положение прямого угла, или даже поднималось еще выше под влиянием непрерывно поддерживаемого сокращения сгибателей предплечья.
Здесь, таким образом, дело идет не только о суммировании раздражений, но и о суммировании самого рефлекторного эффекта под влиянием многократно производимого раздражения сухожилия m. biceps.
Но особенно резко выраженный эффект суммарного рефлекторного эффекта в виде тетанического сокращения мышц руки и в том числе дельтовидной мышцы я наблюдал в некоторых случаях гемиплегии при многократном вызывании описанного мною в свое время акромиального рефлекса путем многократного постукивания молоточком по области acromion. Ввиду особого интереса такого рода явлений я приведу здесь в кратком изложении один ИЗ'случаев, наблюдавшийся мною несколько лет тому назад.
Дело идет о молодом Лице, в возрасте около 25 лет перенесшем несколько лет назад lues и получившем вследствие бывшего с иим инсульта правосторонний гемипарез с своеобразными явлениями ночной слепоты. Потери сознания во время инсульта не было. Больной мог ходить, при-
20*
307
храмывая на правую ногу и слегка волоча носком; правая рука представляется ослабленной, двигать ею больной может, но производит лишь ограниченные движения. Правая половина лииа слегка опущена. Язык не отклонен. В речи расстройств не имеется, но больной не может петь правильно по нотам, хотя заученное ранее поет превосходно и может сам написать ноты для спетого им. Слух с обеих сторон сохранен. Цифры читает также с трудом. Все остальное читает удовлетворительно. При исследовании рефлексов было констатировано ясное повышение коленного и локтевых рефлексов с правой стороны. Состояние болезни до времени исследования в течение нескольких месяцев не представляло существенных перемен, последовало лишь некоторое ослабление параличных явлений. Подробное исследование рефлексов показало, что у больного обнаруживается резкое нарастание рефлекторной возбудимости преимущественно в правой руке. Особенно демонстративно это нарастание обнаруживается при исследовании акромиального и лопаточно-плечевого рефлексов. Если мы будем испытывать акромиальный рефлекс на той и другой стороне, как обыкновенно, одиночными ударами молотка, то мы убедимся, что на правой стороне он получается хорошо, представляясь резко повышенным, тогда как на левой стороне этого рефлекса мы обнаружить не можем, что не представляет какого-либо патологического явления ввиду относительного непостоянства этого рефлекса. Но если мы на правой стороне будем исследовать тот же рефлекс ие однократными ударами молоточка по acromion, а многократным поколачиванием, то повышенный акромиальный рефлекс тотчас же быстро нарастает до той степени, что рука больного постепенно поднимается все выше и выше и доходит до уровня значительно выше плеча вследствие тонического сокращения главным образом m. deltoideus. В конце концов конечность может принять столь приподнятое положение, что предплечье сваливается на голову.
Совершенно аналогичное явление, хотя и в менее резко выраженной степени, можно обнаружить и при исследовании путем многократного поколачивания лопаточно-плечевого рефлекса. Далее, если мы будем подобным же образом исследовать локтевые рефлексы на biceps и triceps, то и здесь мы можем наблюдать нарастание рефлексов, хотя и в сравнительно умеренной степени. Исследование коленного рефлекса, наряду с повышением, также обнаруживает нарастание его силы при многократном поколачивании, но лишь на короткое время, ие более как по истечении нескольких ударов, после чего возбудимость рефлекса временно ослабевает, но затем вновь начинает нарастать, после чего снова временно ослабевает. Таким образом, если мы будем равномерно методически поколачивать молоточком по коленному сухожилию в течение известного времени, то мы получим то усиление, то ослабление рефлекса, периодически сменяющие друг друга. До развития же контрактуры всей конечности, как это бывает во многих случаях, ранее демонстрированных мною в наших собраниях, и как мы имеем в нашем случае в верхней конечности, здесь дело не доходит. Таким образом, нарастание рефлекторной возбудимости в нашем случае обнаруживается наиболее резким образом в верхней конечности и сравнительно слабо выражено в нижней конечности.
Из других явлений отметим, что в верхней конечности быстрое пассивное сгибание руки в локте происходит с рефлекторным препятствием (в два темпа), причем свободно рука разгибается до размера 60 — 70°, и внезапно на этом пункте замечается ясное противодействие временной приостановкой движения вследствие сокращения m. biceps, после чего разгибание вновь происходит даже беспрепятственно. Точно так же и быстрое пассивное сгибание в локте происходит с препятствием на определенном уровне. Первоначально пассивное сгибание происходит вполне свободно приблизительно до уровня, когда предплечье к плечу подогнется приблизительно до уровня прямого угла, на каковом пункте чувствуется ясное противодействие и остановка движения от сокращения m. triceps, после чего дальнейшее сгибание происходит опять свободно.
308
Подобные же явления, хотя и в более слабой степени, можно наблюдать и при пассивном сгибании или разгибании в колейном суставе правой ноги.
Только что отмеченные явления служат особым признаком церебральных органических параличей, что было отмечено мной уже и раньше.
Наконец, существование органического паралича у нашего больного характеризуется еще и тем, что на правой нижней конечности можно обнаружить ясный рефлекс Бабинского.
Ввиду ряда характерных явлений со стороны рефлексов нет надобности говорить, что в данном случае дело идет об органическом поражении мозга, очевидно, развившемся на люэтической почве.
Но для нас не лишено интереса то обстоятельство, что, ввиду явлений музыкальной афазии, правосторонний паралич у нашего больного должен быть признан корковым с возможным участием подкоркового белого вещества.
Так как ранее я имел возможность привести наблюдения, убеждающие в том, что нарастание рефлекторной возбудимости наблюдается в известных случаях спинномозговых заболеваний, а также при церебральных параличах капсулярного происхождения, то, приняв во внимание только что приведенный случай, мы можем с положительностью заключить, что поражения, лежащие на уровне всего центрального двигательного нерва, начиная от его коры до клеток передних рогов спинного мозга, могут сопутствоваться нарастанием рефлекторной возбудимости в большей или меньшей степени.
Так как вышеуказанное нарастание рефлекторной возбудимости не наблюдается, как показывают мои наблюдения, при функциональных поражениях, то представляется очевидным, что этот феномен в свою очередь представляет важный диагностический признак, дающий возможность в известных случаях отличить органический паралич от функционального. В этом отношении нарастание рефлекторной возбудимости получает тем большее практическое значение, что оно составляет, как я убедился, далеко не редкое явление при церебральных параличах, как и описанное мною ранее возбуждение рефлекторного противодействия при пассивном сгибании и разгибании парализованных конечностей в локтевом сгибе и в коленном суставе.
В заключение заметим, что вышеописанные «нарастающие» рефлексы доказывают, что освободившиеся от влияния церебральных центров нижележащие рефлекторные области подвергаются соответствующим функциональным изменениям не только в отношении степени рефлекторной возбудимости, но и в отношении повышения свойственной всем вообще центрам способности суммировать притекающие к ним возбуждения. Отсюда ясно, что вышележащие и, в частности, корковые центры оказывают тормозящее влияние не только на степень возбудимости, но и на способность суммирования возбуждений, притекающих к центрам.
309
Выводы
1. В 1910 г. П. Мари и Ш. Фуа был описан защитный рефлекс гёНехе de defense, а позднее, в 1912 г., и так называемые рефлексы спинномозгового автоматизма — reflexes d'automatisme me-dullaire. Но те же рефлекторные явления описывались мною значительно раньше, в 1900 г., в работе, помещенной в «Обозрении психиатрии» (№9, 1900) и в 1905 г. в том же«Обозрении» (стр. 771).
2. В первой работе я установил, что при повышенной рефлекторной возбудимости при спинномозговых процессах вместе с «быстрым пассивным сгибанием ноги в колене», при котором разгибатели бедра подвергаются пассивному растяжению, «одновременно и совместно сокращаются как разгибатели бедра, так и икроножные мышцы, как будто бы это была одна группа мышц, выполняющая определенное движение». «Иногда этот рефлекс удается вызывать и тыльным сгибанием стопы, приводящим в пассивное растяжение икроножные мышцы». Таким образом, быстрое пассивное растяжение разгибателей приводит к их сокращению в виде разгибательного рефлекса ноги.
3. Другой, менее постоянный рефлекс, обнаруживающийся в тех же случаях, есть «сгибательный рефлекс ноги», вызываемый лучше всего подошвенным сгибанием стопы, а менее часто простым сжатием бедренных и икроножных мышц рукою или даже смещением коленной чашки вытянутой ноги. И в этом случае сгибатели бедра и стопы действуют совместно. Даже тыльное сгибание бедра и стопы может вызвать аналогичный сгибательный рефлекс в виде так называемого парадоксального сокращения мышц.
4. Третьим рефлексом является приводящий рефлекс ноги. Он получается в некоторых случаях при быстром раздвигании ног при согнутых коленях у больных с поражением спинного мозга над уровнем поясничного утолщения. Позднее, в 1905 г. (в «Обозрении психиатрии», стр. 771), я вновь останавливаюсь на сгибательном рефлексе, обусловленном подошвенным сгибанием стопы и пальцев. Его диагностическое значение должно быть поставлено в ряд с другими патологическими рефлексами стопы, как симптомы Бабинского, Оппенгейма, мой сгибательно-пальцевой рефлекс и др., а также описанное мною нарастание костных рефлексов, вызываемых частым и многократным постукиванием молоточком.
5. Кроме вышеуказанного рефлекса, в работе, опубликованной в 1911 г. («Обозрение психиатрии» и Neurologisches Zent-ralblatt, 1911), я описал еще рефлекс, характеризующийся тем же сгибательным движением ноги с отгибанием к тылу стопы и большого пальца, вызываемых многократным постукиванием врачебным молоточком по тылу стопы.
6. Подобный же рефлекс вызывается и уколами стопы и при. том не в подошву только, но и в тыл стопы и в боковые ее части.
310
7. Вышеуказанные рефлексы как последствие прерывания двигательного пирамидного неврона наблюдается иногда даже в тех случаях, где нет ни клонуса стопы, нет рефлекса Бабин-ского или когда последний едва намечен.
8. Заслуживает внимания разивающийся под влиянием суммирования раздражения рефлекс, вызываемый многократным постукиванием молоточком по тылу стопы, ранее того это нарастание силы рефлексов отмечалось мною по отношению к костным, сухожильным рефлексам, что приобретает вообще известное диагностическое значение.
9. Суммирование раздражения выявляется не только соответствующим рефлекторным эффектом, но и в других случаях и нарастанием самого рефлекторного эффекта, что мы видим в некоторых случаях с крайней резкостью при исследовании акромиаль-ного и лопаточно-плечевого рефлекса, в некоторых случаях геми-плегий до такой степени, что при рефлекторном сокращении мышц руки она приподнимается при многократном постукивании даже выше плеча и падает плетью на голову.
10. Нарастание рефлексов в той или другой мере как патологический симптом наблюдается и при исследовании в иных случаях сухожильных рефлексов у гемиплегиков, миэлитиков, и, между прочим, сюда относится описанное мною же явление, состоящее в том, что при быстром пассивном разгибании и сгибании парализованных конечностей происходит в определенный момент рефлекторное временное мышечное противодействие, вследствие чего и то, и другое происходит в два темпа.
ВРЕМЕННОЕ ОЦЕПЕНЕНИЕ ИЛИ ЗАТОРМАЖИВАНИЕ АКТИВНЫХ ДВИЖЕНИЙ КАК ПОСЛЕДСТВИЕ ПОРАЖЕНИЯ ПОДКОРКОВЫХ МОЗГОВЫХ УЗЛОВ
Еще несколько десятков лет тому назад в клинике душевнобольных Военно-медицинской академии помещался больной с своеобразным двигательным расстройством в виде временного застывания членов. Этот симптом заключался в том, что больной, который мог свободно передвигаться по комнатам, внезапно как бы застывал на некоторое время в той позе, в которой заставал его приступ оцепенения. Это своеобразное явление у больного представлялось настолько стереотипным и своеобразным, что обращало на себя внимание всех вообще окружающих. К сожалению, анамнез этого больного совершенно отсутствовал. Несколько лет спустя в Казанской окружной лечебнице я имел возможность, опять же с неясным анамнезом, наблюдать больную с головными болями, явлениями гипомимии и резко выраженной сонливостью, у которой обнаруживалось временами полное на тот или другой период застывание (затормаживание) всех активных движений. Даже во время еды случалось, что больная внезапно становилась неподвижной на некоторое время, застывая с куском во рту. Много позднее, когда был изучен энцефалити-ческий симптомокомплекс, явилась возможность наблюдать много случаев послеэнцефалитического, а иногда, по анамнестическим данным, и послетифозного заболевания, при котором обнаруживались явления гипомимии, причем, если больных заставить поднять брови, то они застывают в приподнятом положении на то или другое время (доктор Мартеновский); с другой стороны, если таких больных рассмешить, то обнаруживается на некоторое время своеобразное застывание улыбки. У одной девушки, перенесшей брюшной и сыпной тифы, при таком же застывании улыбки при явлениях общей скованности, не исключая и мышц лица, имелась некоторая ригидность членов. Все эти симптомы сами по себе не оставляли сомнения, что дело идет в данном случае о протекшем остром инфекционном заболевании с поражением мозговых узлов. У этой больной, помещавшейся в заведываемом мною Воспитательно-клиническом институте под ближайшим наблюдением доктора Д . И. Пескер, между
31?
прочим, вышеуказанный симптом застывания всего тела во время-' движения обнаруживался в необычно резкой степени. Время от времени, повидимому, без всякого внешнего повода больная останавливалась во время передвижения, как вкопанная, и в таком, положении она оставалась долгое время, пока не укладывали ее-в кровать. При этом члены ее при намерении вывести ее из этого состояния обнаруживали напряжение и тугоподвижность. В другие разы у больной те же явления временной оцепенелости происходили в кровати, и в таком положении она оставалась по нескольку часов, оказывая активное сопротивление всякой попытке вывести ее из оцепенелого состояния. При этом больная обнаруживала во время всего припадка явления мутизма. Когда припадок оканчивается, больная встает с постели, проявляя лишь свои обычные болезненные явления. Подробное наблюдение-этой больной приведено ниже.
Совсем недавно я встретился с другой больной, у которой этот симптом временного оцепенения в отдельных членах во время их движения обнаруживался также в ярком виде, причем все сопутствующие явления указывали на поражение основных мозговых ганглиев, хотя определенных указаний на протекшее эн-цефалитическое заболевание здесь и не имелось. Ввиду того что эти явления не являются тождественными с описанным в литературе брадителеокинезисом, а представляют вполне своеобразный симптом, я и считаю возможным описать его в следующих наблюдениях.
Наблюдение 1. Анамнез этого случая, собранный 26/1X 1926 г., заключается в следующем.
Больная Л. И. К-она родилась в Ставрополе на Кавказе в 1901 г. Отец был алкоголиком, перед смертью ослеп и умер от разрыва сердца, а мать скончалась от паралича и водянки. В 1926 г. периодические тяжелые моральные переживания, а именно: 1) муж находился в тюрьме 2 месяца; 2) вслед за тем семья находилась в тяжёлом материальном положении из-за безработицы в течение 9 месяцев; 3) волнения и вспыльчивость семейного характера на почве ревности к мужу, что продолжается и до сего времени.
В 1923 г. впервые прн ходьбе руки не стали двигаться, дальше началось закатывание глаз, вначале раза два в неделю после нервного волнения, теперь ежедневно почти без всякого нервного волнения, а просто после продолжительной ходьбы или переутомления. С тех пор болезнь протекает, постепенно прогрессируя. При этом власть над своими членами стала теряться, например, поднятая ею самой рука не опускается, асе-тается в приподнятом положении до тех пор, пока она не сосредоточит на этом своего внимания или пока ей не укажут на это другие. Взор ее сосредоточен в одну точку с медленной сменой направления, движения при ходьбе стали медленными; больная не может быстро поворачиваться, корпус клонит назад, когда она стоит, паркинсоническое дрожание рук и ног, делать почти ничего не может, а когда кушает, часто застывает в одной позе; иногда при сильном закатывании глаз появляются боли, что сопровождается истерикой.
Начала лечиться в 1924 г. в Физиотерапевтическом институте под наблюдением доктора Венгерова; принимала вначале 15 полуванн с обертыванием и последовательным окутыванием; после этого приняла еще 40 общих водяных ванн. В 1925 — 1926 г. приняла 40 сеансов франклини-
313
'зации и 46 душей Шарко. Никакого улучшения в состоянии здоровья от указанного леч^.йя ie последовало. Лечение кончила в июле месяце нынешнего года вследствие закрытия института на лето. В 1925 г. была у сестры на Кавказе, где без медицинского наблюдения купалась в течение месяца в холодной воде Кубани и принимала солнечные ванны. Никакого улучшения пребывание на Кавказе также не дало. Болезнь особенно заметно прогрессировала за последние два месяца.
По характеру больная очень себялюбивая, капризная и настойчивая, как и все ее сестры, и очень мнительная. Дома все время нервничает и любит ко всему придираться, отсюда частые нервные расстройства. Очень ревнива и любит мужа безгранично, все время беспокоится о том, чтобы муж ее не разлюбил. Когда мужа нет дома, нервничает и не спит по ночам, с приходом его домой успокаивается и спокойно засыпает. В половом отношении до сего времени страстная. Все время скучает, когда она одна, и любит развлечения, читать не может. Еще когда была она здорова, в моменты ревности и семейных разногласий доходила до того, что ломала 'и рвала вещи, выбегала зимой голая на двор. В последнее время боится остаться одна дома и часто устраивает семейные сцены.
Из объективных симптомов, кроме вышеуказанных явлений, отметим, что зрачки хорошо геагируют на свет, в лице гипомимия, обращает на •себя внимание умеренная ригидность мышечной системы и скованность движений при явлениях общего ритмического дрожания рук. Если больной предложить поднять брови, то они застывают на некоторое время в приподнятом положении, как застывает и улыбка. Коленные рефлексы слегка повышены, но никаких патологических рефлексов (Бабинского, моего и др.), указывающих на поражение пирамидного пути, не обнаруживается. Каталепсии при поднятии рук у больной во время исследования не обнаружено.
Наблюдение 2. Приведу еще случай с подобными же явлениями, как и в предыдущем случае. Больной X., около 26 лет, родители здоровы. В 1916 г. падение с лошади, но утраты ориентировки в окружающем не было и больной тотчас же встал на ноги. В 1920 г. заболевает симптомами инфлюэнцы, за которой, по мнению родных, последовал тиф, тогда как на самом деле большого поднятия температуры не существовало, но в течение многих недель имелось длительное состояние бессонницы, а затем в течение нескольких недель была склонность ко сну. Однажды больной На 7 часов оказался в обморочном состоянии без видимой причины. После перенесенной болезни стали развиваться симптомы общей слабости с явлениями паркинсонизма, причем с течением времени в стопе и голени левой конечности временами стала обнаруживаться во время ходьбы судорожная оцепенелость с подгибанием стопы внутрь, препятствующая дальнейшему хождению. Общая слабость, сопровождающаяся ясно выраженной утомляемостью, давала повод наблюдавшим врачам признавать существование миа-•стении, но подробное исследование электрическим током не обнаружило никаких данных, говорящих в пользу этого мнения. Так же и исследование на утомляемость при мышечных сокращениях не дало указаний на миастению. Между тем паркинсонизм у больного выражен тугоподвижностью членов, гипомимией и резко выраженными пульсиями. Достаточно лишь несколько потянуть больного кзади, как он, пятясь, валится назад, также и толчок вперед, а оттягивание за руку в сторону приводит больного к движению в сторону до падения. Во всех движениях обнаруживается резко выраженная брадикинезйя. При поднятии бровей оказывается, что левая бровь недостаточно поднимается, тогда как правая застывает на более или менее долгое время в приподнятом положении, речь слабая, монотонная. Кроме того, совершенно ясно выраженный симптом Кернига в ногах. Сухожильные рефлексы хорошо выражены, мышечные заметно повышены. Дрожания мышц почти нет, за исключением некоторого дрожания языка. Зрачки равномерны, глазных симптомов не имеется. Из двигательных явлений обнаруживает-
•314
ся своеобразный, недавно описанный мною симптом при передвижении, состоящий во временном появлении мелких шажков. Когда больному приходится начинать ходьбу, когда он вынужден поворачиваться или переходить через порог, он временно останавливается и начинает семенить ногами, делая мелкие шажки, после чего он продолжает итти обыкновенным шагом. Бывающая иногда судорожная оцепенелость левой стопы не имеет никакой прямой связи с этим своеобразным расстройством ходьбы. Заметим, что означенный симптом у нашего больного имеет многолетнюю давность, не подвергаясь существенным изменениям.
Наблюдение 3. Больная девочка Ф., 11 лет, была принята в Воспитательно-клинический институт в июне 1921 г. с явлениями в виде припадков сонливости и пароксизмами своеобразной выдыхательной одышки, случающимися по нескольку раз в день. Больная характеризуется педагогами как девочка нервная, раздражительная, легко возбуждающаяся, причем всякое противоречие ее раздражает, и она начинает драться.В ее поведении отмечено, что обыкновенно тихая и вялая она вдруг оживлялась, начинала драться и бить детей меньше ее. За занятиями большую часть времени она дремала и не раз падала со скамейки в период засыпания. Однажды изрезала работу, оставленную возле нее, видимо не отдавая себе в том отчета. В анамнезе отмечается, что со стороны семьи нет указаний на нервные или душевные болезни. В детстве перенесла корь, скарлатину. В 1919 г. перенесла брюшной и сыпной тиф. Последний протекал очень тяжело, с осложнениями. После него девочка сделалась раздражительной, лишилась сна, появились приступы тяжелого дыхания и общей раздражительности. В период последней набрасывалась иногда без всякой причины на окружающих, царапала их и избивала или забивалась в угол, тяжело дыша и не разговаривая.
Объективное исследование показало следующее: больная среднего роста, умеренного питания, вес 1 пуд 35 фунтов, телосложение правильное, кожные и слизистые бледны, грудные железы умеренно развиты, довольно обильная волосистость на лобке, подмышками умеренная, высокое небо, широкие зубы, выстояние верхней челюсти, щитовидная железа в средней доле не прощупывается, а только в боковых долях, небольшая общая ригидность, движения головы назад ограничены, вызывают головокружение и ретропульсию. Кроме того, обращает на себя внимание своеобразная поза больной с приподнятием плеч и приведением их вперед, благодаря чему спина в плечевом поясе кажется округленной, лицо с явлениями гипомимии, чаще всего с полузастывшей улыбкой и полуоткрытым ртом. При покойном положении и опущенных руках обнаруживается дрожание рук и частью ног, больше справа, с паркинсоническими движениями пальцев. Зрачки без существенных изменений в смысле ширины и реакции на свет, но аккомодация вялая. При повороте глаз вправо непостоянный нистагм. На правом глазу симптом Мебиуса. Диплопия. Недостаточность trochlearis. Язык дрожит с незначительной асимметрией, причем правая половина слегка слабее развита. Коленные и ахилловы рефлексы повышены, справа сильнее нежели слева. Ни Бабинского, пи моего рефлекса не имеется. Все виды чувствительности сохранены. Мышечная сила рук и ног без существенных изменений. Симптом Ромберга отсутствует. Походка без изменений. Нет ни антеропульсии, ни ретропуль-сии. Речь с носовым оттенком, смех громкий, отрывочный. Временами повышенное слюнотечение и частое отплевывание. Внутренние органы без существенных изменений.
Во время пребывания в учреждении с июля 1921 г. обнаруживались первоначально плохой сон, отчуждаемость от других детей, раздражительность. Пароксизмами бывает одышка с шумным выдыханием. В это время девочка очень раздражительна и набрасывается на других детей при малейшем поводе. Вместе с тем стали обнаруживаться временами состояния, когда девочка закатывает глаза кверху, приостанав-ливаетсяна месте и остается в таком положении
315
неподвижной, ие реагируя на окружающих и ие отвечая на вопросы.
К концу 1922 г. раздражительность и состояние одышки ослабели,. но стали замечать, что девочка подолгу лежит в кровати и при всякой попытке поднять ее обнаруживалось с ее стороны сопротивление с характером негативизма. В 1923 г. стали вновь обнаруживаться припадки всле-дующем виде: внезапно девочка становится неподвижной, взгляд устремляется в пространства ивтаком состоянии она остается без движения вт. ечение некоторого времени, причем зрачковый и конъюнктивный рефлексы сохранены и надавливанием на подложечную область в одном из первых припадков удалось вызвать живую общую реакцию.
Постепенно эти припадки стали учащаться и удлиняться до 2 —4 часов. Застигнутая припадком в постели, больная сохраняет позу, в которой оиа находилась в момент начала припадка. В таком положении она лежит неподвижно с открытыми-глазами, веки при этом моргают, глазные яблоки отклонены вправо и представляют нистагмообразные движения. Все члены находятся в состоянии, резкой ригидности. Иногда во время припадка она повернет голову и застонет, пожалуется на головную боль, отвернется и снова примет ту же позу. После припадка бывают жалобы на головную боль, но никакого воспоминания о припадке, повидимому, не сохраняет. Большая сонливость вие припадка, общая вялость и малоподвижность, неустойчивость настроения, легкая возбудимость и упрямство. Приемы тиреоидина, повидимому, ослабили припадки, но сонливое состояние осталось попрежнему. С начала 1925 г. припадки вновь учащаются, вне же припадков больная спит по. целым дням. Когда же будят, плачет, прося оставить ее в покое. Одно время припадки повторялись приблизительно по одному в 6 — 7 дней продолжительностью от 2 до 8 часов. Затем припадки становятся, хотя и продолжительнее, — от 5 до 12 часов, но более редкими, повторяясь от 2 до-3 раз в месяц.
Во время лета 1925 г. припадки стали редкими, но сонливость продолжается. Кроме того, больная временами дурашливо произносит разные непонятные слова и выражения, часто плюется, проявляет различные странности, неуместно смеется, хрустит пальцами, плачет, когда пытаются-ее поднять с постели. Со временем саливация и отплевывание прекратились. При явлениях негативизма замечается повышенная внушаемость. Так, больная, встав с постели, пошла в уборную, но там ее застал припадок. Она в с я одеревенела, голова придвинута к груди,, руки приподняты, пальцы коснулись рта, ивтаком положении она оставалась неподвижной. Ее уложили в кровать, в которой она приняла то же положение. После соответствующего внушения в течение х/2 часа мышцы расслабли и конечностям можно было уже пассивно придавать любое положение. Тем не менее, последующие внушения не оказывали на общее состояние больной никакого влияния.
После такого припадка на другой день девочка встала довольная, что ей назначено новое лечение в виде ионофореза Calci chloratii.
Улучшения от ионофореза, однако, не замечается. «Столбняки» снова частые и продолжительные, один из них, например, длился с 3V2 до И часов вечера; 2/V отмечен столбняк в стоячем положении с яйцом в руке. Девочка остановилась на несколько минут, как бы окоченевшая, с прижатыми кгруди. руками. Постепенно голова ее склонялась вперед и, когда лицо достигло горизонтального положения, девочка стала терять равновесие. Ее уложили, в кровать. Глаза были подняты кверху, обнаруживался вертикальный нистагм, хотя на просьбу «посмотреть в глаза» она реагирует и несколько-раз взглядывает на врача, но ригидность мышц остается прежняя.
С июня 1926 г. припадки снова стали реже, сонливость также улучшилась, до обеда уже не спит, а часто и после обеда. В поведении неряшлива
316
я проявляет резкий негативизм при попытках остановить от того или другого несоответствующего действия. Припадки по 6 — 7 в месяц. Пьет вообще довольно много, особенно после впрыскивания triplex и применения лечебных ванн.
21/VIII 1926г. девочка в улучшенном состоянии переведена в вспомогательную школу.
Не касаясь других интересных особенностей данного случая, мы привели его только с целью иллюстрации временного парок-сизмального затормаживания всех активных движений с проявлениями ригидности членов и негативизма у больной с явлениями паркинсонизма.
Итак, у всех наших больных, наряду с другими явлениями, указывавшими на поражения основных мозговых ганглиев, обнаруживался своеобразный симптом временного затормаживания движений и застывания членов и даже всего тела при активных движениях. Этот симптом следует отличать от каталептических явлений, нередко наблюдаемых у больных, страдающих энцефалитическими и послеэнцефалитическими явлениями, ибо каталепсия характеризуется сохранением положения при пассивном смещении членов, тогда как застывание тела или членов в определенном положении наблюдается при активных движениях, и притом совершенно неожиданно для больных, ибо это застывание застает больных иногда в неудобной позе, которую они выдерживают подолгу, после чего это застывание самостоятельно прекращается. В том случае, когда застывание проявляется в отдельных движениях, например, в застывании поднятых бровей, в застывании улыбки и в застывании отдельной конечности во время ее движения, это застывание обыкновенно прекращается с тем вместе как обратить внимание больного на это застывание, которое тогда под влиянием активного сосредоточения и прекращается.
Дело идет в этих случаях, очевидно, о совершенно особом и своеобразном симптоме, более или менее полном, причина которого еще неясна и который заслуживает специального изучения. Есть все основания полагать, что дело идет здесь о временном возбуждении внепирамидной системы проводников, которая, временно сковывая активно движущиеся члены, приводит их на время в состояние оцепенелости. Играет ли в этом случае какую-либо роль вегетативная (парасимпатическая и симпатическая) нервная система, выяснят дальнейшие наблюдения.
Выводы
1. У больных с явлениями паркинсонизма и вообще явлениями, связанными с поражением мозговых узлов, обнаруживается иногда своеобразный симптом временного затормаживания членов или даже всего тела в определенном положении на то или другое время в периоды активных движений.
317
2. Этот симптом следует отличать от каталепсии, наблюдаемой нередко при той же болезни в форме удерживания членов в том или ином положении при пассивных смещениях их, а не при активных движениях членов тела.
3. Вышеуказанное затормаживание движений, развиваясь внезапно, застает больных в той или другой, часто даже неудобной, позе, которую они выдерживают подолгу, после чего оно мало-помалу ослабляется само собой или же прекращается вместе с тем, как обращалось внимание больных на это застывшее, своеобразное положение членов больных.
4. Патогенез симптома пока неясен. Но возможно полагать, что здесь дело идет о временном возбуждении внепирамидной. системы проводников.
IV ТРУДЫ ПО ПСИХИАТРИИ]
СЛУЧАЙ ПОЛНОЙ СЛЕПОТЫ, СОПРОВОЖДАЕМОЙ ГАЛЛЮЦИНАЦИЯМИ ЗРЕНИЯ
(Протоколы заседаний общества психиатров в Петербурге 10 декабря 1883 г.)
Д-р В. М. Бехтерев демонстрировал случай полной слепоты, сопровождаемой галлюцинациями зрения.
Больная около 30 лет от роду. В половине июня прошлого года у нее без особенной причины развились сильные стреляющие боли в задней части головы, сопровождающиеся головокружением. Вместе с этим начало развиваться ослабление зрения на обоих глазах. Головные боли в особенности были сильны в течение первых двух недель, затем они ослабели, но и до сих пор все еще появляются приступами в сопровождении головокружения. Ослабление зрения возрастало до сентября, когда оно достигло степени полной слепоты на оба глаза. Около того же времени начали появляться двигательные расстройства в виде нарушения равновесия и наклонности отклоняться при движении влево; наконец, присоединились зрительные галлюцинации. Ни сифилиса, ни травматических повреждений головы у больной не было.
Status praesens. Полная слепота на оба глаза, причем отсутствует даже количественное восприятие света. Несмотря на это, оба зрачка, хотя и представляются расширенными, но еще вполне ясно сокращаются под влиянием сильного света; на акко-модативные импульсы, когда больную заставляют направить глазные оси на кончик своего носа, зрачки также ясно сокращаются. В обоих глазах замечается резкий нистагм. Офталмоско-пически — обоюдосторонний застойный сосок. Из двигательных расстройств обращает на себя внимание только то обстоятельство, что больная слегка колеблется на ногах; это колебание еще немного усиливается с закрытием глаз.Кроме того, при ходьбе больная отклоняется в левую сторону, после же нескольких пассивных или активных вращений тела она почти не в состоянии удерживаться на ногах. Из субъективных симптомов, кроме временно появляющихся головных болей, больная жалуется на чувство головокружения, сопровождающееся чувствомр пово-
21 В. М. Бехтерев
321
рачивания головы и туловища вправо. Галлюцинации зрения,, продолжающиеся i до настоящего времени, состоят в самых разнообразных видениях. Вначале перед ее глазами мерцали искры и светлые круги; она видела движущихся перед нею животных (собак и лошадей), затем женщин и детей в разноцветных платьях, наконец, целые пейзажи: улицу, покрытую снегом, по которой двигаются сани, блеск луны на небе и т. п. Во время приступов головокружения предметы галлюцинаций нередко двигаются в одном направлении, большей частью слева направо, т. е. в направлении, противоположном субъективному или кажущемуся движению ее тела. Ни в чувствительной, ни в двигательной сфере нет резких изменений. На основании всех существующих явлений д-р Бехтерев допускает, что в этом случае дело идет о поражении четверохолмия, причем патологический процесс захватил также и подлежащие волокна, связывающие центральное серое вещество третьего желудочка с мозжечком. По мнению В. М. Бехтерева, сообщенное наблюдение приобретает особенный интерес в отношении того, что при слепоте, зависящей от поражения четверохолмия, во-первых, еще возможна реакция зрачков на свет, а во-вторых, могут появляться сложные зрительные галлюцинации. Последний факт, таким образом, может служить веским доводом против теорий, по которым происхождение галлюцинаций обязано возбуждению инфракорти-кальных центров (в данном случае — четверохолмия) или органов перцепции, по выражению некоторых авторов. Кроме того, В. М. Бехтерев указывает на то интересное обстоятельство, что головокружение, испытываемое больной, отражается на локализации ее галлюцинаций во внешнем пространстве.
О ЛЕЧЕНИИ НАВЯЗЧИВЫХ ИДЕЙ ГИПНОТИЧЕСКИМИ
ВНУШЕНИЯМИ
Не очень давно я напечатал клиническую лекцию, в которой изложил применяемый мною способ лечения навязчивых идей самовнушениями в начальных периодах гипноза и привел тяжелый случай навязчивых идей, кончившийся выздоровлением, благодаря применению только что сказанного лечения1. Между прочим, в этой лекции я упомянул, что «в течение нескольких последних лет в своем курсе психиатрии при изложении учения о навязчивых идеях я всегда обращал внимание слушателей, между прочим, и на применение послегипнотических внушений с целью лечения навязчивых идей».
В то время, когда я писал эти слова, мне еще не удалось провести на практике лечение навязчивых идей послегипноти-ческими внушениями, так как у наблюдавшихся мною больных с навязчивыми идеями хотя и могли быть вызываемы самые начальные явления гипноза, но ни один из больных не мог быть приведен в более глубокие степени последнего, в которых, как известно, лучше всего удаются послегипнотические внушения.
С тех пор, однако, мне представилось несколько случаев с вышеуказанным поражением, в которых можно было вызывать степени гипноза, вполне позволявшие применение с лечебными целями и гипнотических внушений. При этом я вполне убедился в могущественном влиянии гипнотических внушений на навязчивые идеи. Последние при всем своем упорстве заметно ослабевали уже после первого же сеанса и в благоприятных случаях окончательно исчезали после нескольких сеансов; в менее же благоприятных случаях лечение хотя и затягивалось на более продолжительный срок, но все же рано или поздно приводило к благоприятному исходу. При этом успех лечения обусловливался собственно не только степенью развития, продолжительностью и упорством навязчивых идей, но и тем обстоятельством,
1 В. М. Бехтерев, Случай навязчивых идей, излеченный самовнушениями в начальных периодах гипноза (клиническая лекция), Вестник клинической и судебной психиатрии и невропатологии, 1890, т. VII, вып. 2.
21*
323
какая степень гипноза могла быть вызвана у больного, в зависимости от чего, без сомнения, изменялось и действие гипнотических внушений. Так как собирание подходящего материала в указанном направлении продолжается мною и по сие время, то я пока и не имею в виду приводить здесь все случаи навязчивых идей, пользованные мною гипнотическими внушениями, и ограничусь лишь одним наблюдением, которое прекрасно поясняет целебное значение гипнотических внушений по отношению к навязчивым идеям.
К-, замужняя, 40 лет, дочь очень нервной матери и отца, злоупотреблявшего спиртными напитками. Две сестры ее отличались нервностью. Сама больная в молодости была нервной и в девицах страдала бледною немочью. Года 4 спустя после выхода замуж, т. е. лет 18 тому назад, у нее впервые обнаружились припадки большой истерии, бывшие, впрочем, всего раза два или три. Но с тех пор время от времени случаются припадки малой истерии, выражающиеся давлением в горле, сердцебиением и слезами, а изредка и хохотом. Кроме того, больная с давних пор обнаруживает чисто болезненную боязнь некоторых животных, например, кошек и мышей, боязнь привидений, а иногда и боязнь нечистоты, в силу которой она принуждена часто мыть свои руки. Вместе с тем она поразительно нерешительна во всем и тревожится по всяким пустякам. Поводом к тревогам нередко служат даже такие маловажные события, как посылка куда-нибудь прислуги с самым обыденным поручением. Наконец, по временам у больной бывали и более резкие навязчивые идеи; так, если она бывала в обществе, то ей нередко казалось, будто она обидела то или иное лицо своим разговором. Эти мысли об обиде кого-либо обыкновенно подолгу преследовали больную и нередко приводили ее в сильное волнение. Но особенно резко состояние ее ухудшилось недели за три до первого ее визита ко мне в конце декабря. Совершенно неожиданно с ней случилось следующее происшествие, послужившее поводом к развитию мучительных навязчивых идей: она посоветовала одной из своих родственниц, г-же М., страдавшей уже лет 16 чахоткой, поехать на поклонение мощам святого Германа в Свияжск. Г-жаМ., действительно, послушалась ее совета, но случилось так, что приехав из Казани в Свияжск, она умерла как раз у цели своего путешествия, в самой церкви. Известие об этом сильно поразило нашу больную. С тех пор ее стала преследовать мысль, что ее родственница умерла благодаря ее неуместному совету. Несмотря на внутреннее разубеждение самой больной, эта мысль с того времени не оставляла ее в покое почти ни одной минуты: она часто и подолгу оплакивала как свою родственницу, так и свой поступок. Беспокоившие ее ранее мысли, вечные сомнения, тревоги и различные страхи теперь отошли на второй план и как бы перестали существовать для больной. Ни развлечения, ни занятия ее уже не успокаивают. Будучи в обществе, она не слышит, что говорят кругом ее, и все думает об одном и том же; хозяйство тоже не идет ей в голову вследствие той же причины. Даже ночью больная нередко видит свою несчастную родственницу в сновидениях; вместе с тем ей часто представляются те или другие события, касавшиеся смерти последней. Поэтому больная спит в высшей степени беспокойно, часто просыпается, тревожась и ночью той же мыслью.
При объективном исследовании (17 января) найдено следующее. Больная несколько ниже среднего роста, правильного телосложения, с довольно бледными покровами, но достаточно развитым подкожным жировым слоем. Легкие в порядке. В сердце замечается лишь некоторая возбудимость и наклонность к учащеннрй деятельности. Со стороны кишечника можно отметить некоторую вздутость. Никаких других расстройств внутренних органов не замечается. Чувствительность и двигательная сфера, а равно и рефлексы без существенных изменений.
324
Назначив больной бромистый натрий, который, впрочем, она и раньше принимала без особенного успеха, я предложил ей, кроме того, лечение гипнозом, на которое она решилась не без колебаний. При первой попытке гипнотизирования больная, наслышавшаяся много о гипнозе, начала обнаруживать сильное волнение; появились даже признаки предстоящего истерического припадка. Ввиду этого, я воспользовался самой слабой степенью сна, применив на первый рал описанный мною ранее способ самовнушения. Как только больная сомкнула веки под влиянием пассов, я заставил ее повторять за мною следующие слова: «отныне я не должна более тревожиться смертью М., потому что она умерла не вследствие моего совета, а вследствие того, что была больна уже давно и могла бы одинаково умереть и дома точно таким же образом». Последствием этого самовнушения было то, что уже на следующий день (18 января) больная чувствовала себя лучше, не плакала совсем и хорошо спала ночью. Навязчивая мысль, по ее словам, как бы отдалилась от нее. Правда, она еще приходит на ум, но уже не так беспокоит больную и скоро исчезает из сознания. Дальнейшим последствием самовнушения было то, что больная теперь относилась к гипнозу уже без всякого волнения и могла быть, действительно, в короткое время загипнотизирована с помощью пассов, причем во время гипноза ей было внушено не мучиться более мыслью о смерти г-жи М. Явления гипноза заключались в том, что больная не могла произвольно открыть глаз; вместе с тем у нее появлялось резкое притупление чувствительности к болевым раздражениям и некоторое расслабление членов.
Для испытания силы внушения в этом относительно легком гипнотическом состоянии я приказал больной сжать в кулак кисть правой руки, заявив после того, что она не может более расправить пальцы; и, действительно, больная некоторое время вовсе не могла раскрыть кулака, а затем хотя и раскрыла его, но лишь после долгих усилий. Следует, впрочем, заметить, что подобные же внушения, хотя и в значительно меньшей степени, удаются у больной и в бодрственном состоянии.
На следующий после гипноза день (19 января) больная заявила, что после вчерашних внушений весь день до вечера чувствовала себя хорошо, но с 8 часов вечера вновь появилась мысль о смерти г-жи М., по ее мнению, вследствие того, что она целый вечер оставалась одна. Ночью в сновидении больная тоже видела М., но утром чувствовала себя довольно покойно. Следует заметить, что со вчерашнего дня у больной открылись месячные, во время которых прежде она всегда чувствовала себя несколько хуже и страдала сильнее навязчивыми идеями. Гипноз вызывается теперь заметно глубже прежнего. Внушения сделаны те же.
20 января больная, явившись ко мне на прием, заявила, что весь вчерашний день после гипноза чувствовала себя очень хорошо и навязчивые идеи в течение дня'совсем не появлялись. Ночью спала с 11 до 3 часов, затем проснулась и некоторое время тревожилась прежней мыслью о г-же М., к 5 часам заснула, однако хотя и видела снова во сне М., но, проснувшись утром, чувствовала себя так же хорошо, как и с вечера. Так как больная должна была дней на пять уехать домой в деревню, то, загипнотизировав ее, я внушил ей, чтобы во время своей отлучки из Казани она не думала вовсе о г-же М., и чувствовала себя также и ночью покойно, не видя вовсе М. и во сне. Гипнотический сон у больной теперь получался уже глубже прежнего, и сила внушений вследствие того изменилась: теперь у больной в состоянии гипноза повторными внушениями можно было вызвать параличи, сведения и полную анестезию, что прежде не удавалась.
Уехав после 20 января, больная вернулась ко мне лишь 12 марта с заявлением, что прежние мысли о г-же М. у ней совершенно исчезли вслед за последним сеансом. По ее словам, от прежнего у ней осталось лишь нервное состояние; но зато в последнее время больную вновь стали беспокоить явления, бывавшие у нее до навязчивой идеи о смерти г-жи М. Так, ее смущает поразительная нерешительность во всем: если, например, ей нужно сделать какое-либо распоряжение по хозяйств-у, то она постоянно
325
сомневается, как правильнее поступить, и долго не может на что-нибудь решиться; решившись же, меняет свои распоряжения раза по три. Кроме того, она попрежнему стала бояться некоторых животных (кошек, мышей) возвратилась также и боязнь привидений и нечистоты. В силу последней она постоянно мост руки, иногда бессчетное число раз в сутки; часто даже моется и вся раза по три в день.
Несмотря на усиленные просьбы больной начать немедленно же гипнотическое лечение для избавления ее от только что перечисленных явлений, я должен был, за недостатком времени, отклонить ее просьбу до ближайшего времени, назначив ей микстуру из 5JJJ бромистого калия, 3J3 антипирина и зjj лавровишневой воды на 5vj перегнанной воды по 6 столовых ложек в сутки.
В течение следующих 5 дней больная чувствовала себя, однако, во всех отношениях хуже, отчасти, может быть, под влиянием месячных. Сомнения беспокоили ее уже до того, что она не могла написать даже и простого письма без многочисленных колебаний. Ей казалось, что она все перепутала, написала не то, что следует, и потому должна была даже просить других лиц прочесть ее письма для проверки. Под влиянием тех же сомнений больная по многу раз запечатывает и распечатывает свое письмо, прежде чем решается его послать.
Вместе с тем все ей кажется нечистым: платье, руки и даже то, что она ест; при этом она испытывает нередко безотчетный страх по самому незначительному поводу; так, если ей нужно пойти в другую комнату, то она боится отправиться туда одна; ей кажется, что там кто-нибудь стоит и может ее напугать. Сверх того, по временам больная подвергается сильной реактивной тоске.
Ввиду такого состояния больной я решил уже не медлить гипнотическим лечением и 17 марта, загипнотизировав ее, внушил, чтобы она более не мучилась сомнениями, не тосковала и вообще не испытывала никакой боязни, между прочим, и боязни нечистоты.
Через день, 19 марта, больная сообщила мне, что сомнения ее уже не беспокоят в такой мере, как прежде; боязнь нечистоты меньше, и вообще она чувствует себя лучше. Снова сделано внушение в состоянии гипноза.
21 марта, прибыв на прием, больная заявила, что последние 2 дня она чувствовала себя сравнительно очень хорошо; и если прежние яления еще и обнаруживаются, то лишь в весьма слабой степени. Снова сделано внушение в гипнозе и опять с благоприятным результатом.
Оправившись совершенно от мучивших ее навязчивых идей, больная уехала к себе в деревню, где остается и до настоящего времени, не чувствуя потребности в дальнейшем лечении.
Итак, в первый раз, в январе, потребовалось всего 4 гипнотических сеанса (из которых один с самовнушением), чтобы освободиться от мучительной навязчивой мысли, а второй раз, в марте, было достаточно трех гипнотических сеансов, чтобы устранить из сознания больной целый ряд навязчивых идей.
Результат этот, безусловно, говорит сам за себя и не нуждается в особых пояснениях. Во всяком случае способ лечения навязчивых идей гипнозом, на мой взгляд, заслуживает самого серьезного внимания со стороны врачей-специалистов.
О СЛЫШАНИИ СОБСТВЕННЫХ МЫСЛЕЙ
В последнее время в психиатрической литературе, благодаря сообщению д-ра Кеппена х было обращено внимание на своеобразный симптом при душевных болезнях, который может быть назван слышанием собственных мыслей.
Этот симптом, известный уже и ранее, представляет, на мой взгляд, совершенно особый интерес, в силу чего он уже давно служил предметом моего внимания. Еще более 7 лет тому назад, во время моей профессорской деятельности в Казанском университете, я посвятил клиническую лекцию одному из больных, у которого вышеуказанный симптом обнаруживался в крайне резкой степени. Этот больной, бывший чиновник, человек в общем довольно интеллигентный, представляет собой старого алкоголика, пившего водку в продолжение 35 лет и только в 1886 г. совершенно оставившего эту пагубную привычку. Он сделал, между прочим, собственное описание своих обманов чувств, из которого я извлеку прежде всего все существенное, относящееся к его болезненному состоянию.
«Я житель Самарской губернии, был 25 лет на государственной службе, с 1872 г. проживал в своем небольшом имении, занимаясь сельским хозяйством; но хозяйство расстроилось и имение продано в 1875 г. В этот -год, проживая несколько месяцев в Самаре один без семьи, я заболел от последствий алкоголизма и, главное, к ужасу своему почувствовал, что при мне находится какое-то существо, благодаря которому в левом ухе как будто бы раздается металлический звонок; последний, не давая мне покоя ясным голосовым звуком, но как будто издалека, говорил о моем дурном семейном положении и производил в то же время страдание действием на нервы всего тела, причем я чувствовал то подергивание нервов, то осыпание тела сверху огненными искрами, . отчего я приходил в состояние беспокойства. В таком виде я поступил в самарскую больницу и, не получивши облегчения, оставил оную через месяц.
Из Самары я переехал в Саратов искать службу, но существо, живущее при мне, или, вернее сказать, во мне, не оставляло меня. Я вновь пользовался у одного из докторов и был в городской больнице, но лекарства (мушка и морфий в малых, вероятно, дозах) мало действовали на меня, разговор со мною и возбуждение не прекращалось, я спал не более двух часов в сутки и в то же время не чувствовал усталости.
От нервного ли расстройства, или от беспрерывных действий на меня непонятного существа на всем теле начал высыпать сплошной лишай.
1 К б р р е п, Ueber das Gedankenlautwerden, сообщение в Обществе психиатров и невропатологов в Берлине 19 марта 1889 г.
327
В таком виде я из Саратова по своим делам отправился в Москву, где, пользуясь с месяц лекарствами из лечебниц исключительно от лишая, наконец, должен был лечь в Екатерининскую больницу. При этом ясный человеческий разговор в левое ухо не прекращался, когда же не было говора то я чувствовал, как и теперь чувствую, в левом ухе как бы металличе ский тончайший звонок. Здесь начало повторяться осыпание меня искрами еще сильнее, я становился беспокойным в ночное время, проводил ночи без сна, и меня едва не отправили в дом для душевнобольных, но оставили, потому что ответы мон были ясны и толковы и притом я, скрепя себя, не стал объяснять доктору, что со мною происходит. По осмотре меня в больнице профессором Найденовым, я пользовался ваннами и мазями от открывшейся на мне лишайной сыпи и, по совершенном почти очищении тела, через пять недель вышел из больницы. Четыре месяца после того я жил на даче под Москвой, ведя самую строгую жизнь в родном семействе, но ни нравственная жизнь, ни физическая сила не помогли мне в том, чтобы я освободился от находящегося во мне существа, которое мучает меня своим вмешательством как в одиночестве, так и в обществе.
Из Москвы я возвратился в Саратов, потом жил в Самаре, Оренбурге и в каждом из этих городов был в больницах, но бесполезно. Высказывая врачам откровенно свое положение, я вызывал в них лишь сомнение в здравости своего рассудка, выносил долговременные мушки без всякой надобности, причем врачи большей частью смотрели на мое состояние, как на последствие невоздержной жизни и желание притом отдохнуть в больнице, ввиду ограниченности моих ничтожных средств (пенсион в 100 руб. в год).
По моим понятиям, существующее во мне не могло появиться от моей натуры, потому что нет ничего в нем согласного и сходного с моими правилами жизни; у него постоянная ложь и обман, вообще отсутствие всего нравственного и постоянные скверные ругательства. Это что-то чуждое, прививное. Оно смотрит на предметы отчетливо, указывает моему вниманию предметы, которых я не расположен даже и видеть, и может даже отвлекать от предмета, затемняя самое зрение. Всех лиц, которых когда-либо я знавал или даже где-либо встречал, оно указывает уже издали, называя фамилии даже и в том случае, если бы я не досмотрел или не хотел обратить внимания. Где я предположил быть или предполагал что-либо сделать полезного, оно старается отвлекать меня, затемняя зрение, наводит другие мысли и, наконец, им производится физическая боль в руках и ногах, а при письме трясение рук или онемение самих пальцев, так что с трудом могу пересилить это действие.
От действий существующего во мне я хожу в возбужденном состоянии ежедневно, иногда вдруг налагается тяжесть на меня, чувствую, что верхнее платье на мне неестественно по своей тяжести, не дает мне хода, едва даже могу войти в дом к кому нужно по делу, при этом трудно объясняюсь, глаза затуманиваются и упорно отворачиваются от лица, с кем приходится объясняться. Нередко оно останавливает ход мыслей, по его выражению, „наводит столбняк", и тут же обращается с вопросом: „О чем Вы, Николай Иванович, думаете теперь?", оно же закрывает иногда слух так, что я нередко не разбираю, что говорят со мною, и не могу читать на близком расстоянии. Зрение от натуры моей всегда было хорошее, а с появлением во мне непонятного существа при чтении я прибегаю к очкам, а равно и при письме, иногда даже через очки не могу разбирать печатного, при усиленном же напряжении зрения я достигаю того, что явное противодействие теряет силу. Несколько случаев было таких: вдруг зрение мое очищается, я совершенно ясно и свободно могу читать и писать без очков и в то же время следовал всегда вопрос: ,,Что хорошо, Николай Иванович, с чистыми глазами?" и вскоре затем зрение попрежнему становилось туманным.
До начала августа 1888 г. я два года проживал в Астрахани, вел самую строгую жизнь, не допускал ни в чем излишества, а спиртуозного совершенно не употреблял, развлекаясь исключительно посещением публичной библиотеки и домашним чтением, и тем не менее существо не оставля-
328
ло меня и тяготело надо мною. Ему также доступно чтение. Последнее происходит таким образом: я развертываю книгу и смотрю в нее построчно, при этом идет отчетливое чтение само собой, то одним ясным голосом, то другим; но я не допускаю подобного чтения, потому что оно вообще меня утомляет.
В продолжение четырнадцати почти лет я не имел женщины и не имею потребности, между тем в стволе и мошонке я нередко чувствую боль как будто бы от постороннего действия чем-то острым.
Как ни грустно и тяжело мое положение столько лет, я не могу, однако, плакать, слез нет, чувства сердечные связаны.
Пятый уже месяц я проживаю в Казани с целью обратиться к пособию медицины и именно в клинике и заходил в нее неоднократно, но во мне делается какое-то отупение и охлаждение к своему положению и я опять возвращаюсь в квартиру, между тем действия существа и по сие время не прекращаются в различных видах, которые трудно описать в подробности; но, главное, и теперь продолжается металлический как будто бы шум в левом ухе, иногда над самой головой, а также наколы как бы иглой по разным частям тела, в особенности когда я стараюсь днем или ночью заснуть и, лишь совершенно измучившись, засыпаю часа на три и много на четыре.
Все описанное — краткий обрис всего происходящего со мною по настоящее время; для меня жизнь мучительна и безотрадна и если берегу ее и берег до сего времени, то из покорности к провидению и в надежде, что это явление, если еще небывалое в медицинской практике, разъяснится на пользу других».
В дополнение вышесказанного на лекции от 4 февраля 1889 г. больной сообщил, что он болен «мышлением», сам он будто бы мыслить не может, так как каждый раз, когда он начинает мыслить, то все его мысли тотчас же произносятся вперед; он старается переменить ход мыслей и вновь за него мыслят вперед. Этот разговор он слышит всегда левым ухом. Голос, который больной слышит уже в продолжение 13 лет и притом всегда в левое ухо, вполне ясный, мужской, но с различным характером — то более тонкий, то более грубый, а иногда сиповатый. Этот голос всегда слышится больным на близком расстоянии как будто бы кто-нибудь говорит около его левого уха. По утверждению голоса, левое ухо — его (голоса); а правое ухо— больного. Иногда больной испытывает около уха как бы движение под кожей чего-то постороннего. Иногда он чувствует там же ощущение как бы от укуса блохи или пчелы. Нередко также около его левого уха появляются ощущения ползания постороннего существа (наподобие чувства ползания клопа), а также и своеобразные ощущения в самом ухе. В последнем часто слышится шум, «как будто в ухе сидит пчелиный рой». Если больной заткнет правое ухо, то шум еще сильнее. Все вместе взятое и заставляет его думать ,что в его левом ухе или около последнего поселилось какое-то существо, могущее говорить человеческим голосом, которого он, однако, никогда не видит. Это-то существо и овладело его мыслями. Оно подсказывает вперед его собственные мысли, иногда же говорит ему то, с чем он не согласен и против чего он готов протестовать всей
329
душой или, наконец, делает различные замечания по поводу того или другого его поступка или его слова, даже прямо противоречит больному и вступает с ними в спор. Если больной стоит в церкви, то о н слышит нередко поющий голос, который вперед напевает нее то, что поют на к л и р о с е. Но иногда голос в церкви путает его мысли, говорит дурное, даже произносит площадную брань. Однажды в соборе в Москве во время эктеньи больной пришел в ужас от того, что услышал, и должен был выйти из церкви. Вообще в церкви больному нередко слышатся неприятные слова, но он старается от них уклониться и путем некоторой борьбы ему удается заставить голос затихнуть.
Если больной идет по улице и видит, например, вывеску, то голос вперед его читает то, что написано на вывеске (например, фамилию портного, сапожника и пр.). По словам больного, это не он читает, так как он иногда даже не думает о вывеске и, несмотря на то, голос прочитывает ее содержание. Если больной завидит издали кого-либо из знакомых, то г о л о с тотчас жео бы кновен-н о еще прежде, чем он подумает об этом лице говорит ему: «вот идет такой-то». Иногда больной даже и не думает обращать внимание на проходящего, а голос своим заявлением.по поводу проходящего невольно заставляет его обратить на него внимание. В другой раз его внимание отвлекается от известного предмета и он его не видит, хотя бы предмет лежал перед ним. Это явление больной называет затемнением зрения, которое является лишь на известные предметы. Например, больной нуждается в очках и не может их видеть, хотя бы они лежали перед глазами, а должен нащупать их рукой. По его заявлению, это ничуть не зависит от его невнимания, так как он следит за собой.
Когда больной читает книгу, то чтение повторяется его голосом; мыслить в это время он не может, если он останавливается, то притихает и голос. Если больной, не читая, смотрит глазами в книгу, то ему читает книгу голос. Без очков, например, он не может читать, но если без очков он смотрит в книгу, то его глазами как бы кто-то смотрит и читает. Впрочем, он убедился, что существо, которое ему говорит в левое ухо, нуждается в очках так же, как и он сам.
В начале появления голоса больной был поражен таким явлением, он даже бегал от голоса, не знал куда деваться, как будто его кто-то преследовал. «Есть что-то такое в натуре человека, — замечает больной, — чего, может быть, до сих пор еще не объяснили себе ученые. Этот голос не может быть только испорченностью слуха; нет ли в организме существ каких-нибудь?» задает он себе вопрос. В силу этого больной и допускает существование особого существа, которое говорит ему в левое ухо.
330
Иногда голос говорит обыденные вещи: например, после посещения больным меня голос говорит: «Наконец-то занялись Вами, Вы не лечились в больницах; напрасно в клинику идете». Почему «напрасно» — голос не сказал. «Вы, говорит, смерти не боитесь», на что больной заметил, что он хладнокровен.
По заявлению больного, ему очень трудно становится управлять собой: ему вперед уже говорят, куда идти, иногда туда, куда он и не хочет; он делает напротив и является борьба в мыслях и действиях. Благодаря этому, он испытывает постоянное напряжение. Существо может заставить больного заснуть тогда, когда ему нужно идти, и наоборот; в последнем случае является возбуждение и больной может пройти очень много без усталости. Как производится существом это возбуждение, он не знает, но, во всяком случае, существо может заставить приказать делать то. что е м у у г о д-н о. Если больной хочет уснуть, оно ему часто не дает сна и глаза остаются открытыми. «Хочу закрыть глаза, закрываю — и должен их открыть по приказанию голоса, потому-что становится тяжело. Хочу п и с а т ь, б е р у перо — пальцы раздвигаются, писать не могу, если голос этого не ж е л а е т». Т о же и с ногами: больной идет, а его ноги раздвигаются в стороны; ему становится трудно идти и напряженно сдвигать ноги. Иногда его глазами двигают в ту сторону, к уд а он не ж е-л а е т. Нередко ноги в том или другом месте колют как бы жесткой щеткой, и он берет сам щетку и трет это место, после чего колотье переходит в другое место. Под влиянием того же существа воображение рисует больному чрезвычайно живые картины. Прежде он допускал в воображении эти картины, но они его утомляли; теперь же, когда появляются картины, он отвертывается к лампе, и тогда картины пропадают. Картины являются различные: знакомые местности, люди и пр. Иногда бывают и незнакомые картины. Больной сам может вызвать свои картины в ответ на картины существа; это он может сделать каждый раз при открытых и закрытых глазах. Иногда больной слыхал гнилостные, трупые запахи.
По поводу причины всех этих явлений больной задается вопросом: «Все ли в натуре человека исследовано?» Он не допускает существования большого существа, а — маленького; может быть не в ухе а в голове, может быть за ухом, в ухе, по всему организму. На вопрос, как может меленькое существо говорить человеческим голосом, больной отвечает: «Не управляет ли это существо слухом?» Он может определенно сказать, что существо это есть
331
и управляет его организмом. Это не есть результат болезни, так как он слышит голос очень ясно и притом столько лет; в то же время он рассудок имеет, не потерял его. «Ведь в медицине много неизведанного!» — добавляет больной.
Когда я высказал больному убеждение, что это галлюцинации и что я могу ему представить более полутораста человек, имеющихся в Окружной лечебнице, которые слышат голоса, тотчас же больной заметил, что «может быть они слышат не так ясно!» На дальнейшее разубеждение больной заметил, что он, подобно Галилею, должен стоять на своем и утверждает, что «все же есть существо».
Объективное исследование ушей, произведенное проф. Высоцким, показало следующее: сильная сухость слуховых проходов, особенно левого; мутность левой барабанной перепонки и резкое общее Втягивание ее, отсутствие светового конуса. Ослабление звуковой проводимости через воздух на левое ухо и усиленный резонанс на камертон в этом же ухе через головные кости, что вместе взятое указывает на усиленное внутриушное давление. Звук часов слева едва слышит даже при непосредственном приложении. Костная проводимость звука, однако, вполне сохранена. На правое ухо тоже имеется ослабление, но в значительно меньшей степени; часы слышит в расстоянии более одной четверти от уха. В зрачках резкой разницы нет. Язык обнаруживает дрожание и небольшое отклонение в сторону. Укол булавкой в различных частях вызывает лишь приятное чувство, а не боль; сухожильные рефлексы заметно повышены.
В пополнение вышеприведенной истории болезни необходимо заметить, что все вообще явления у больного оставались без изменений в течение дальнейшего наблюдения, продолжавшегося около года, за исключением лишь того, что больной со временем стал уклоняться от рассуждений на тему о существе, живущем у него в левом ухе и даже стал отказываться от этой идеи, но, повидимому, неискренно; все же галлюцинаторные явления и также слышание вперед своих мыслей у больного продолжались в прежней степени. Между прочим, ему была сделана попытка ослабить явления путем гипноза, но, к сожалению, сон больного оказался очень слабым и внушение имело лишь кратковременное действие. Больной в течение первого вечера после гипнотизации, пока вспоминал слова внушения, не слышал повторения своих мыслей и его голос умолкал; больной также спал всю ночь превосходно, как никогда в течение болезни, но па другой день голос опять беспокоил больного попрежнему.
К сожалению, больной после нескольких раз гипнотизации прекратил свои посещения и, таким образом, нельзя было выяснить ее влияние на болезненное состояние. Между прочим, в течение сеансов гипнотизации выяснилось, что даже непродолжительная фиксация блестящего предмета вызывает у больного зрительные г а л-
332
люцинации. В первый раз во время фиксации блестящего шарика больной заявил, что он ясно видел двое часов со стрелками. В другой же раз больной при смотрении на блестящий шарик видел уже сильно жестикулировавшего человека, который наклонялся и как будто что-то искал; при фиксации на близкое расстояние он увеличивался, при смотрении вдаль он уменьшался. Кроме того, больной видел мужчину с детьми, которые находились при нем. При этом голос говорил те или другие фразы, например, указывая на мужчину с детьми, повторял: <Это ты!».
Если мы обратимся теперь к выяснению болезненного состояния, то не может подлежать сомнению, что мы имеем здесь дело с галлюцинациями алкогольного происхождения, составляющими остаток неизлеченного галлюцинаторного помешательства, проявлявшегося в прежнее время гораздо более бурными симптомами и вместе с тем более распространенными галлюцинаторными явлениями, которые вместе с затиханием болезненного расстройства ограничились почти исключительно левым ухом и обусловили своеобразный бред. Упорное появление галлюцинаций в левом ухе, очевидно, обусловливается хроническим ка-тарральным состоянием среднего уха при общей возбудимости центральных органов. Так как катарр у больного ограничивался почти исключительно левым ухом, то этим легко объясняется тот факт, что галлюцинации приняли одностронний характер. Не касаясь подробнее этой интересной особенности нашего больного, мы остановимся с самого начала лишь на том явлении, которое может быть названо слышанием собственных мыслей.
Мы уже упомянули, что симптом этот недавно обратил на себя внимание в психиатрической литературе, благодаря сообщению д-ра Кеппена в Берлинском обществе психиатров и невропатологов. Больной д-ра Кеппена, страдавший хронической пара-ноей, в августе 1892 г. встретил в ресторане даму, которая взглянула на него странным образом. По возвращении домой он впервые услышал женский голос, который повторял его собственные мысли, критикуя их ьслух. С тех пор больной стал слышать как различные голоса, главным же образом женский голос, повторяют и критикуют вслух его собственные мысли. Если, например, больной что-нибудь пишет, он слышит как кто-то произносит вслух написанное. Особенно беспокоит это явление больного в то время, когда он не занят; при усиленных же занятиях явление это вообще ослабевает и даже прекращается совершенно. Автор обращает внимание на то, что его больной при этом никогда не ощущал движения языка и губ.
При обсуждении вышеуказанного симптома Кегшен относит его к слуховым галлюцинациям, которые он разделяет на две группы: одни не имеют ничего общего с обычным порядком мышления больного и другие, стоящие в тесной связи с мыслями больного, который галлюцинирует образами, имеющими ближайшее отношение к предмету его идей.
ззз
Первая группа галлюцинаций объясняется центральным или периферическим раздражением органов чувств, тогда как вторая группа галлюцинаций, к которым должно быть отнесено и своеобразное явление, состоящее в слышании собственных мыслей, должно быть объяснено иначе. По взгляду Мейнерта, в этом случае дело заключается не в раздражении органов чувств, а в извращенном толковании своих мыслей или мозговых процессов. Между тем, Крамер слышание собственных мыслей объяснял раздражением мышечного аппарата речи. Последний взгляд Кеппен совершенно справедливо оспаривает. По его мнению, если бы причина этого симптома состояла в раздражении мышечного аппарата речи, то больные в этом случае должны были бы полагать, что кто-нибудь приводит в движение их язык, губы и пр. или что они сами принуждены двигать языком или губами, как это, действительно, и наблюдается при так называемых двигательных галлюцинациях. Между тем ничего подобного на самом деле у больных такого рода не наблюдается.
По мнению д-ра Кеппена, сущность явления состоит в расстройстве слуха. В нормальном состоянии каждое мысленное слово звучит акустически, вызывая иннервационное ощущение выговаривания или произношения этого слова. По Кеппену, этот-то процесс ложно истолковывается больными, в особенности параноиками, в силу чего они полагают, что посторонние люди произносят вслух их мысли.
На мой взгляд, не подлежит никакому сомнению, что слышание собственных мыслей должно быть отнесено к порядку галлюцинаторных явлений. Так, мой больной, как можно видеть из описания его болезни, сохранил еще хороший интеллект и способен отлично анализировать себя; при этом он решительно отри- • цает, чтобы его языком кто-нибудь управлял, тогда как он ясно сознает, что вся суть дела заключается в слышании голоса, в громком повторении этим голосом его мыслей при письме, при чтении, а также в тех или других замечаниях и критике его мыслей и действий со стороны этого голоса, который больной, по крайней мере первоначально, приписывал особому существу.
Нельзя, однако, согласиться и с мнением Кеппена, что слышание собственных мыслей основано на ложном истолковании слухового звучания слов. Дело в том, что в нашем случае собственно дело шло не только о повторении мыслей больного, но и о произношении их вперед, как, например, при чтении или при встрече знакомого. Иногда и другие мысли больного как бы подсказывались вперед. Это подсказывание мыслей, как известно, у параноиков составляет вообще нередкое явление, а между тем, вряд ли можно полагать какое-либо различие по существу между тем, что понимается под слышанием собственных мыслей и так называемым подсказыванием их, состоящим в сущности также в слышании собственных мыслей, но слышании вперед.
334
На мой взгляд, в этом случае дело идет не о том, что слуховой отзвук мысленных слов ложно истолковывается, а о том, что этот звук, благодаря необычайной возбудимости центрального аппарата, усиливается в такой степени, что, будучи аперцепи-рован, он достигает интенсивности объективно произносимого слова.
Если внимание больного сосредоточивается на своих мыслях, то слуховой отзвук, превращающийся в галлюцинаторный образ, аперцепируется вслед за аперцепцией мыслей больного и тогда больной слышит лишь повторение собственных мыслей, наоборот, если больной сосредоточивает свое внимание на громком слуховом отзвуке, то он аперцепируется раньше его мыслей и больной слышит слуховой отзвук своих мыслей вперед, следовательно, имеет так называемое подсказывание мыслей. Фигурально представляя самый процесс, можно его уподобить тому, как дело происходит в часах Вундта с восприятием одновременных слуховых и зрительных впечатлений. Если мы сосредоточиваем внимание на слуховом впечатлении, оно слышится вперед; если, наоборот, мы сосредоточиваем внимание на зрительном впечатлении, мы его видим ранее слухового впечатления. Разница заключается лишь в том, что в этом случае дело идет о произвольном или активном внимании, тогда как в состояниях болезни мы имеем дело не только с активным, но и с пассивным вниманием, которое привлекается к ходу мыслей или к слуховому их отзвуку, помимо личности больного.
О ВНУШАЮЩЕМ ВЛИЯНИИ СЛУХОВЫХ ОБМАНОВ ЧУВСТВ
Уже многими авторами было обращено внимание на тот факт, что душевнобольные с сохранением интеллекта допускают в своем бреде такие нелепости, которые никоим образом не могут быть совместимы с более или менее здравой логикой.
Это особенно рельефно выразилось и в случае, описанном нами в работе о слышании собственных мыслей. Больной наш, как мы видели, человек крайне интеллигентный и в болезни рассуждающий вполне здраво по всем вообще предметам, не относящимся к его бреду, утверждает тем не менее положительную и очевидную для всех и каждого нелепость, что в его левом ухе или около уха поселилось какое-то невидимое существо, которое говорит человеческим голосом и управляет всем его организмом. Надо заметить, что у нашего больного бред ограничивался почти одной этой идеей и в то же время в период наблюдения у него был поражен галлюцинациями почти один только орган слуха, к тому же с одной лишь стороны, и при всем том больной, несмотря на свой интеллект, не мог отрешиться от этой идеи. В противовес здравым взглядам в невозможности такого существа, он довольствуется одними предположениями, догадками или вопросами, не дающими ему возможности окончательно отрешиться от этой идеи.
Заканчивая свою записку, он, между прочим, выражает надежду, что его признание послужит в пользу науке и, если дело идет о явлении, еще небывалом в медицинской практике, наивно замечает больной, что оно разъяснится на пользу других. В другом месте по поводу своего существа больной восклицает: «Мало ли в медицине есть неизведанного?». Таким образом, несмотря на то, что больной не находит возможным опереться в своей логике на что-либо положительное, кроме фантастических предположений и наивных вопросов, он твердо держится своей болезненной идеи и после моих настойчивых убеждений, сделанных в присутствии целой аудитории, в ошибочности его взглядов, он тем не менее сослался на Галилея, заявив, что, как последний утверждал, что земля вертится, так и он должен стоять на своем, т. е. что существо в его левом ухе действительно существует.
336
Обыкновенно говорят, что упорство бредовых идей, обусловленных обманами чувств, основывается на том, что последние в глазах больного являются свидетельствами органов чувств гораздо более убедительными, чем все убеждения других. Это обстоятельство, действительно, в некоторых случаях имеетогром-ное и безусловное значение, в особенности же там, где галлюцинации необычайно живы, обильны и обнаруживаются во многих органах чувств, благодаря чему для больного вместе с убедительностью галлюцинаторных образов утрачивается и всякая возможность правильного контроля со стороны непораженных органов чувств. В нашем случае, однако, не было ничего подобного: галлюцинации, как уже упомянуто, сосредоточивались главным образом в одном из органов чувств, именно, в органе слуха и притом лишь на одной стороне; к тому же больной имел полную возможность убедиться в болезненном происхождении своих слуховых галлюцинаций прежде всего потому, что в его левом ухе имеется шум, свист и в то же время обнаруживается очевидная слабость слуха, испытываются ненормальные кожные ощущения в наружном ухе и в частях, соседних с ушной раковиной, и пр. Словом, у больного существовал целый ряд болезненных явлений, понятных всем и каждому, не говоря о возможности более или менее правильного контроля со стороны всех остальных органов чувств, и, несмотря на то, он твердо убежден в справедливости своей нелепой идеи о присутствии в его левом ухе особого существа.
Такое упорство идеи, несмотря на очевидную ее нелепость, вполне напоминает собой внушенные идеи. Последние, как известно, также прочно удерживаются в сознании в течение известного времени, несмотря на очевидное противоречие их с здравой логикой.
Повидимому, сила внушенной идеи основывается главным образом на том, что внушение входит в сферу сознания совершенно независимо от «я» субъекта, которое поэтому может оказывать лишь слабое воздействие на этот прививной продукт сознания.
Не то же ли самое мы имеем и в галлюцинациях? Последние, являясь продуктами бессознательной умственной деятельности, точно так же вступают в сознательную сферу, как нечто совершенно независимое от «я» субъекта и потому-то они в известных случаях с такой быстротой подчиняют сознание больного даже наперекор его логике и здравому смыслу, который в этом случае легко и быстро вступает в ту или другую сделку с новым гостем сознания. Словом, на мой взгляд, одной из причин подавляющего влияния галлюцинаций на сознание душевнобольных является то обстоятельство, что их галлюцинаторные образы происходят из бессознательной сферы, уже в силу того быстро подчиняют сознание и волю подобно тому, как внушенные идеи, появляясь в сознании, подчиняют последнее, несмотря
22 В. М. Бехтерев
337
даже на то, что они не совместимы с здравой логикой и убеждениями данного лица.
Самый характер галлюцинаций в подобных случаях напоминает собой настоящие внушения: наш больной, например, однажды слышал почти постоянное твержение на следующую тему: «Не ходи в клинику, нас не найдут и не поймут, а ты все равно умрешь. Да ты не боишься смерти, а мы-то куда? Мы ведь тоже существа» и пр.
Очевидно, что слышание таких фраз действует подобно внушению и прививает больному идеи, которые при других условиях легко могли бы быть исправлены сознанием больного.
Известно уже давно, какое магическое влияние производят на больных отрывочные слуховые галлюцинации с повелительным тоном («убей!», «подожги!»), которые нередко сразу овладевают сознанием больного и приводят иногда тотчас же к преступным деяниям. В подобных случаях внушающий характер обманом чувств ясен до очевидности. В других случаях больные сами отлично сознают свою неспособность противостоять подчиняющему влиянию слуховых обманов чувств и говорят о внушении им чужих мыслей.
В нашем случае внушающее влияние обманов чувств проявлялось не только в отношении развития бреда, но они, по словам-больного, вызывали остановку мышления, а также действовали на физическую сферу, вызывая, например, затемнение зрения по отношению к известным предметам, совершенно напоминающее собой явление, известное в гипнозе под названием отрицательной галлюцинации, или же вызывая особые изменения в напряжении мышц нижних конечностей, затрудняющие ходьбу больного, и производя трясение рук при письме или онемение пальцев. Что дело идет в этом случае о настоящем внушении со стороны-обманов чувств, производимом в бодрственном состоянии, легко доказывается тем обстоятельством, что каждый раз временное затемнение зрения или те или другие двигательные расстройства и расстройства чувствительности предшествуют слышанью голосов, предупреждающих о наступлении соответствующих явлений.
Особенно нередко у больных такого рода наблюдается влияние слуховых галлюцинаций на болевые ощущения, которые в полном смысле слова внушаются предшествующими им голосами, предупреждающими о предстоящем вызывании этих болевых ошущений.
ОБ ИСКУССТВЕННОМ ВЫЗЫВАНИИ
ОБМАНОВ ЧУВСТВ У АЛКОГОЛИКОВ,
СТРАДАЮЩИХ ГАЛЛЮЦИНАТОРНЫМИ,
ФОРМАМИ ПОМЕШАТЕЛЬСТВА
В последнее время Липман х обратил внимание на возможность легко вызывать зрительные галлюцинации и иллюзии у больных, страдающих запойным бредом, с помощью надавливания на глазные яблоки. При этом выяснилось, что у некоторых]; больных возможность вызывать такие иллюзии и галлюцинации наблюдалась и вслед за прекращением острого периода запойного бреда. По наблюдениям автора, такие искусственно вызванные обманы чувств отличаются от естественных тем, что предметом их содержания редко служат животные, как, например, крысы или мыши; напротив того, в них в большинстве случаев преобладают неодушевленные предметы или люди;при этом не обнаруживалось и устрашающего характера искусственно вызванных обманов чувств^ которые представлялись обыкновенно лишенными связности и того колебательного или качательного движения, которое, по автору, столь характерно для естественных галлюцинаций запойного бреда. По мнению автора, его способ может быть полезным и как диагностический прием в тех случаях, когда больные отрицают присутствие обманов чувств.
В видениях, вызываемых давлением на глазные яблоки, автор видит, между прочим, точное доказательство этиологической зависимости галлюцинаций от периферического раздражения2.
На основании своих опытов он согласен также принять, что многие из самопроизвольных обманов чувств, которые не представляют собой иллюзорного восприятия внешних впечатлений, являются иллюзорной переработкой внутренних, т. е. эндо-органных, возбуждений. Автор считает даже вероятным предположение, что такова причина Есех вообще обманов чувств запойных пьяниц.
Нужно заметить, что уже и ранее некоторые Еысказьшались в пользу зависимости обманов чувств при запойном бреде от
1 Arch.i . Psych., Bd. XXVII, 1.
2 Ztl. f. Nervenheilkunde, Oktobrr 1895.
339
22*
периферических рэздражений в органах чувств. Не говоря о старых наблюдениях Жолли, вызывавшего у галлюцинирующих больных слуховые обманы чувств с помощью раздражения слухового органа постоянным током \ и наблюдениях Кеппена, описавшего появление обманоЁ чувств при введении воронки в ухо и при других аналогичных манипуляциях2 специально по отношению к больным, страдающим delirium tremens, Некке3 допускает, что иллюзии зрения у них происходят вследствие энтоптических световых явлений. Независимо от вызываемых гиперемией сетчатки искр, огненных шариков и т. п., по Некке, легко может •случиться, что глазное яблоко придет в соприкосновение с краем постели и таким образом произойдет периферически обусловленное сильное световое явление, которое затем соответственным образом перерабатывается в больном мозгу.
Вышеуказанная работа Липмана, впрочем, не осталась без возражений. Д-р Альцгеймер4 убедился, что зрительные галлюцинации вышеуказанным способом могут быть вызваны у разнообразных больных, как, например, параноиков, истеричных, эпилептиков и даже у паралитиков, в силу чего Альцгеймер не находит в этом явлении ничего специфического или характерного для запойного бреда. Тем не менее Липман и после этого заявления, не отрицая фактов, сообщенных Альцгеймером, допускает известное диагностическое значение искусственно вызываемых зрительных обманов, хотя и далек от того, чтобы считать их явлением специфическим5.
При своих наблюдениях над алкогольными формами помешательства я уже в течение многих лет обращал особенное внимание на искусственное вызывание обманов чувств, но способ, к которому я прибегал для этой цели, был совершенно иным. Для вызывания слуховых иллюзий и галлюцинаций я пользовался монотонным звуком молоточка от индукционного аппарата, к которому больные должны былиприсл'ушиваться, для вызывания же зрительных обманов чувств я прибегал к фиксации больными в течение известного времени блестящего предмета, находящегося вблизи глазных яблок, как это применяется при гипнотизации 6. И тем, и другим способом мне удавалось вызывать в высшей степени легко обманы чувств не только у алкоголиков, находящих-
1 По Жолли, в этих случаях дело сводится, собственно, к раздраже-мию п. trigemini, которое рефлекторно передается на центры слуха (Beitr. Z. Theorie d. Halluzinationen, Arch. f. Psych., 1874, Bd. IV).
2 К δ p p e π, Gehirnstorungen u. Psychose, Allg. Zschr. f. Psych., Bd. XXIV.
3 N а с k e, Beitrage z. Lehre d. Delirium tremens, Dtsch. Arch, f. Min. Med., 1880, Bd. XXV.
4 Zbl. f. Nervenheilk., Oktober 1895.
5 Zbl. f. Nervenheilk., Marz 1896.
8 He менее удобным способом вызывания слуховых обманов чувств может служить, по всей вероятности, всякий продолжительно, однообразно звучащий инструмент, как электрический камертон и т. п.
340
ся еще в периоде запойного бреда, но также и спустя более или менее продолжительное время после исчезновения обманов чувств и притом в некоторых случаях с поразительным постоянством. Я имел больных, у которых после перенесенной белой горячки не оставалось уже ни одного из болезненных проявлений и тем не менее достаточно было больному несколько прислушаться к звуку индукционного аппарата, как он уже начинал слышать в этом звуке голос, произносящий те или другие слова. Равным образом и при фиксации блестящего предмета у таких больных легко вызывались те или другие видения, например, какие-либо предметы, физиономии людей и т. п. Такого рода явления со стороны слухового и зрительного органа у больных, перенесших delirium tremens, продолжаются различное время; иногда они исчезают уже спустя несколько дней по прекращении бреда, в других же случаях наблюдаются целыми неделями и даже в течение нескольких месяцев. Наконец, в случаях, когда после перенесенного delirium tremens обманы чувств у алкоголиков остаются как постоянное явление, нередко остается навсегда и эта поразительная наклонность к появлению обманов чувств при вышеуказанных условиях в органе слуха или зрения или же в одном из этих органов.
Прекрасным примером подобного состояния может служить больной, на котором мы остановились при рассмотрении вопроса «О слышании собственных мыслей» г. Мы видели, что у этого больного, перенесшего много лет назад приступ острого галлюцинаторного психоза алкогольного происхождения и у которого остались галлюцинации, ограничивающиеся почти исключительно одним левым ухом, уже при непродолжительной фиксации блестящего предмета в первый раз появилось видение двойных часов; в другой раз при такой же фиксации блестящего предмета больной уже видел сильно жестикулировавшего человека, затем мужчину с детьми и пр. Совершенно аналогичные явления при одинаковых условиях наблюдались и в других аналогичных случаях.
Точно так же прислушивание к монотонному дребезжанию индукционного аппарата у таких больных уже по истечении короткого времени вызывает появление слуховых обманов. В некоторых случаях прислушивание ко всякому решительно звуку, на котором может быть сосредоточено внимание больного, уже вызывает появление слуховой иллюзии того или иного содержания.
Относительно характера обманов чувств, вызываемых у алкоголиков искусственно с помощью вышеуказанных раздражений, необходимо заметить, что, согласно мнению Липмана, они, действительно, в большинстве случаев не представляют ничего устрашающего и вообще почти не содержат чего-либо неприятного для больных в противоположность естественным
1 Обозрение психиатрии, 1896, № 10.
341
обманам чувств, обнаруживающимся при алкогольных формах помешательства.
Что касается причины искусственного вызывания обманов чувств при вышеуказанных способах, то оно, повидимому, сводится не на одну только повышенную возбудимость самих органов чувств, но, вероятно, еще в большей мере на влияние напряженного внимания. Дело в том, что вышеуказанные раздражения оказываются действительными обыкновенно лишь в том случае, если больные сосредоточивают на них внимание, в противном случае те же самые раздражения неспособны вызывать обманов чувств.
Таким образом, очевидно, что какое бы значение в происхождении обманов чувств мы ни придавали внутренним раздражениям периферических органов, не подлежит сомнению, что в появлении обманов чувств в вышеуказанных случаях играет известную роль и чрезмерная возбудимость психических центров, приходящих в состояние патологического возбуждения под влиянием направления внимания на то или другое внешнее впечатление.
Что касается, наконец, вопроса относительно диагностического значения искусственно вызываемых обманов чувств для алкогольных форм помешательства, то, хотя и не может подлежать сомнению, что подобные же явления могут обнаруживаться в известных случаях и при других формах помешательства, тем не менее мои наблюдения говорят с решительностью в пользу того что вышеуказанные явления составляют в общем довольно частый симптом галлюцинаторных форм именно алкогольного происхождения.
О ПЕРИОДИЧЕСКИХ ПРИСТУПАХ РЕТРОАКТИВНОЙ
АМНЕЗИИ
В нижеследующем изложении дело идет о больном, который после постигшего его удара стал обнаруживать кратковременные приступы амнезии. Больной сам описал свое состояние в особой записке, которую я и приведу здесь ранее, чем передать другие сведения о его болезни.
«Болезнь моя заключается в повторяющихся припадках временной потери памяти; во время этих припадков я забываю, что было со мною даже час назад, не помню, какой в это время день, где я был перед тем; иногда забываю, что было накануне; но период самозабвения не простирается дольше суток перед тем моментом, как я очнусь от самозабвения, ибо в последнем я легко припоминаю и вообще помню все то, что было за сутки до наступления припадка потери памяти.
Припадки потери памяти обыкновенно случаются со мною после продолжительных непрерывных служебных занятий, в особенности требующих умственного напряжения.
Пользующий меня врач, хорошо изучивший мою натуру, приписывает эти явления переутомлению, при расстройстве нервной системы, и потому прописывает мие бромистый калий в микстуре в соединении с другими лекарственными веществами, в то же время безусловно запрещая мне служебные занятия. Прием бромистого калия всегда оказывал полезное на меня действие; припадки скоро проходят даже и без приема медикаментов, если я день или два отдохну от занятий, а иногда даже через два-три часа отдыха.
Начало расстройства нервной системы относится к концу 1874 г. В 1873 г. я потерял первую жену, о которой долго и сильно тосковал, оставшись с тремя малолетними детьми. Тоску я старался заглушать усиленными служебными занятиями, даже н на дому. В конце же 1874 г., около 20 декабря, я перенес сильнейшее душевное огорчение, непосредственное последствие которого выразилось тем, что я с 4 часов дня до 9 часов утра следующего дня заснул и так крепко, что никак не могли разбудить меня ни к вечернему чаю того же дня, ии к утреннему на другой день, поэтому на другой день утром пригласили ко мне врача, который велел поставить мне по сторонам позвоночного столба сухие банки, что и было исполнено тогда же, отчего я очнулся. Через несколько дней меня отвезли в Военно-медицинскую академию к приват-доценту Петру Ивановичу Успенскому. Это было в начале января 1875 г., когда уже возобновилось в академии чтение лекций студентам. П. И. Успенский при мне же читал студентам лекцию о моей болезни, сказав, что со мной случился сильный нервный удар, сопровождающийся воспалением мозговой оболочки, вследствие усиленных занятий при недостатке воздуха, и что болезнь впоследствии повторится. Он назначил мне куратора студента и прописал йодистый калий. Лечение продолжалось у меня на квартире около месяца
343
под наблюдением куратора; тогда же начались у меня непроизвольные подергивания мускулов лица и век; у меня изменилось выражение лица и глаз, так что видевшие меня тогда говорили, что я не похож на себя. Физически я стал вскоре поправляться и начал ходить на службу, но заниматься ничем не мог, вследствие того, что при малейшем умственном напряжении появлялась у меня острая, колющая боль в затылке. Летом, того же 1875 г. я уволен был в отпуск на 3 месяца на родину, во Владимирскую губернию. Там я значительно окреп, но выражение лица и глаз оста лось прежнее.
После того у меня сильно развилось сердцебиение, унаследованное от моих родителей и усилившееся после смерти первой моей жены. (В октябре 1875 г. я вторично женился). Сердцебиение продолжается до настоящего времени. Кроме того, я стал слишком впечатлителен, не могу слушать рассказов или чтения о несчастных случаях не только с людьми, но и с животными; тогда я моментально весь вздрагиваю. Это вздрагивание случается у меня и тогда, когда я, легши в постель вечером, о чем-либо думая, долго не засыпаю.
Возвращаюсь к обстоятельствам, следовавшим за указанным выше нервным ударом.
В 1875 г. я лечился электричеством, но бесполезно; мне помогло лечение сгущенным воздухом и пребывание в летнее время на родине, в отпуске. Тогда же я начал по совету доктора каждодневные прогулки пешком в течение часа и более, что продолжаю, хотя и не всегда, до настоящего времени.
С 1877 г., считая свое здоровье восстановившимся, я начал свои служебные занятия, которые с 1878 г. должны были усилиться.
Что касается до нынешних%припадков временной потери памяти, то первый такой припадок случился со мной летом 1890 г., но довольно слабый; более сильные припадки начались в 1894 г. Летом этого года я, живя на даче в Лесном, усиленно занимался одной служебной работой, причинявшей мне под конец головные боли, сначала тупые, какие обыкновенно бывают от утомления, а потом и острые, щемящие, в задней части головы, в левой половине ее; но в этот раз припадка потери памяти не было. По возвращении с дачи в город, осенью 1894 г., у меня усилились служебные занятия по так называемой текущей части; тогда повторился со мной сильный припадок временной потери памяти; то же и той же причины повторилось со мной в начале 1895 г. По причине своего очень заботливого характера, я, засыпая очень поздно, около 12 часов ночи, очень рано просыпался и так как, лежа в постели, долго не мог заснуть, то уже вставал с постели и приступал к занятиям часа в 4 в 3 утра и даже раньше; иногда по окончании работы я, снова успокоенный, ложился на постель; иногда, хотя очень редко и не надолго, опять засыпал. Осенью того же 1895 г. и в начале 1896 г. у меня снова усилились служебные занятия; тогда оба раза снова повторились припадки потери памяти, которым всегда предшествовала головная боль, сначала тупая, а потом острая, щемящая, в левой стороне затылка; в таких случаях я прерывал занятия на некоторое время. Но весной 1896 г. те же болезненные припадки стали повторяться чаще, причем у меня развилась чрезмерная впечатлительность. Мне говорили врачи, что надо бросить служебные занятия, но я не решался ослаблять свои занятия. В то же время я не мог не сознавать и могущие произойти от неумеренного труда вредные и опасные для моего здоровья последствия. По настоянию врачей летом 1896 г. (в конце мая) я взял месячный отпуск и совершил прогулку по Волге в течение 2 недель; с Волги я ездил иа родину дней на 10. Прогулка эта очень хорошо повлияла на здоровье, так что я чувствовал себя вполне здоровым и способным даже к усиленным служебным трудам, которые и продолжал до лета 1897 г.; летом сего года, живя на даче в Шувалове, я очень усиленно занимался одним сложным служебным делом, не оставляя и обыкновенных своих всегдашних деч, для чего каждодневно ездил с дачи на службу. На даче опять повторился припадок потери памяти, но довольно слабый и непро-
344
должительный. Осенью и зимой от той же причины опять стали повторяться эти припадки. В начале февраля текущего года (4-го числа), повторился легкий припадок потерн памяти, а 6-го числа того же месяца он продолжался с 9 часов утра до 6 вечера; в этот промежуток времени я ничего не помнил из недавнего, не теряя, однако, как и прежде, сознания; не ходил и на службу; на другой день я не помнил того, что было со мною накануне. 8-го числа я совершил прогулку за город на извозчике; освежился, но несколько простудился; чувствовал на другой день боль в по-хровах головы; затем легкое ослабление памяти опять повторялось и иа первой неделе великого поста. В феврале месяце я в первый раз был у Вас на квартире в приемные часы вместе со своей женой. Когда Вы расспрашивали меня о моей болезни, я, находясь в тревожном состоянии, даже плакал, что обратило на себя Ваше заботливое внимание. Как ласковое обхождение Ваше со мной, так и прием прописанных Вами лекарств произвели благотворное действие на меня: я стал крепче, головных болей не было, так что я продолжал свои служебные занятия беспрепятственно, хотя их было и много. Поэтому-то во второй внзит к Вам (6 марта) я совершенно справедливо сказал Вам, что стал чувствовать себя гораздо лучше с того дня, как был у Вас. Головной боли у меня теперь не бывает, несмотря на довольно напряженные служебные занятия. Раз у меня повторился весьма легкий н кратковременный припадок потери памяти. Это было, когда я на днях шел из бани, где была очень высокая температура. То же случалось со мной и прежде, уже давненько. Головной боли у меня теперь не бывает, а иногда чувствую только утомление; тогда я оставляю ненадолго занятия, и головные боли прекращаются.
В дополнение к вышеизложенному считаю нужным сказать, что я всегда вел, как и теперь веду, вполне регулярный образ жизни; водку пью более чем умеренно. Редко хожу куда-либо в гости по вечерам, но возвращаюсь домой рано. Повседневно же ложусь в постель в начале 11-го часа, читаю, лежа в постели, газеты или книги, около 12 часов засыпаю. Просыпаюсь, по большей части рано: часов в 6, 7; иногда встаю и занимаюсь служебными делами.
Ввиду всего вышеизложенного, естественно, возникает вопрос: соображения как легкие, так и напряженные относятся к области умственной деятельности; казалось бы, они должны отражаться на уме, на ослаблении умственных способностей. Почему же от умственного переутомления страдает память и почему от переутомления забываются только обстоятельства самого последнего времени, но сохраняются в памяти обстоятельства и события, бывшие прежде, даже в очень отдаленное время. Как объяснить это физиологически?».
В дополнение к этому описанию я прибавлю от себя лишь то, что служит к большому уяснению состояния нашего больного и опишу болезненные явления, наблюдавшиеся позднее, 16 марта 1898 г., к каковому времени относится вышеприведенная записка больного.
Первый приступ с больным случился в 1890 г. и продолжался несколько часов, затем приступов не было вплоть до 1894 г., когда они снова стали изредка повторяться. Прошедшим летом случился приступ такого же рода, длившийся всего один час. Затем с 31 января нынешнего года до половины марта месяца было уже 3 приступа. Первый приступ 31 января имел всего продолжительность около получаса, а в начале февраля приступ длился с 8 1/2 часов утра до 9 часов вечера. Последний приступ случился с больным вечером 15 февраля и продолжался всего полчаса. Затем с больным случилось еще два приступа в период-
34Г,
моего наблюдения за ним, о которых речь будет ниже. Все приступы, которые е ним бывали раньше, развивались после напряженных умственных занятий; им обыкновенно предшествуют головные боли, особенно в затылочной области. Развиваются эти приступы всегда внезапно и также внезапно кончаются. Во время приступов у больного вполне сохраняется сознание относительно всего окружающего, он сознает свое болезненное состояние, сильно удручается им, иногда даже плачет, сознавая ненормальность своего состояния, описывает его окружающим и пр. При этом никаких болезненных явлений со стороны двигательной сферы не обнаруживается.
Сущность самого психического состояния, как это явствует из его многократных описаний, делаемых во время самых приступов, состоит в том, что он при полном сохранении сознания теряет память по отношению к событиям недавним, предшествовавшим лишь за несколько часов до приступа и никак не более суток. Все же более отдаленные события он отлично припоминает и рассуждает вообще здраво. Его обычные в этом случае жалобы заключаются в заявлениях о невозможности припомнить ничего из того, что он делал ранее за несколько часов, куда ходил, с кем говорил и т. п. Больной даже удивляется тому, что он не может припомнить ничего из того, что было так недавно, тогда как отдаленные события помнит хорошо, как всегда. После того как приступы проходят, у больного развивается обыкновенно по отношению к ним амнезия и он потом большей частью не может припомнить о состоянии, бывшем с ним, или же припоминает смутно, неясно. Последние два приступа, случившиеся в марте месяце, произошли с больным первый в жаркой бане, второй под влиянием тревоги; но оба приступа протекли совершенно стереотипно, как и все предыдущие. Ни головокружений, ни судорожных явлений у больного во время приступов вообще никогда не обнаруживалось. К анамнестическим сведениям необходимо добавить, что после бывшего с ним удара у него наблюдалась некоторое время impotentia virilis, затем половая функция восстановилась. Сифилисом и другими венерическими болезнями он никогда не страдал.
Что касается объективного исследования, то его результаты сводятся к следующему: больному в настоящее время 67 лет, он высокого роста, хорошего телосложения, имеет довольно высокое служебное положение по комитету раненых и представляет собой человека вполне интеллигентного. В сосудах обнаруживается склероз, во внутренних органах заметных отклонений от нормы не имеется. У больного обнаруживается не совсем внятная речь, что идет еще от времени удара. Пишет левой рукой, вследствие того, что в правой руке с 60-х годов развилась mogigraphia, но после бывшего удара и левой стал писать плохо, как он выражается, «крючками». Теперь он может писать правой рукой лишь с сильно поднятым локтем и сильно отведенным большим паль-
316
нем. При всем том правой рукой долго писать не может,вследствие развивающейся судороги пальцев. В обоих глазах staphiloma posticum, в левом глазу очень большая v. d. = 0,3, v. sin. = 0,2. На дне глаза видимых изменений нет.
При исследовании мочи, произведенном в химической лаборатории д-ра Пеля, найдено: суточное количество 1 100, реакция кислая, удельный вес 1,023. В осадке мочи, кроме слизи и мостилыюго эпителия, найдены кристаллы мочевой кислоты, лейкоциты, семенные тельца, гиалиновые и мелкозернистые цилиндры. Мочевины 27,43 г, мочевой кислоты 0,88 г, хлористого натрия 12,43 г, фосфорной кислоты 2,07 г, серной кислоты 1,96 г, общее количество азота 14,59 г, левкомаинов 1,23. Кроме того, следы белка и следы пептонов. По расчету отношение общего количества азота мочи к количеству азота мочевины выражается 13,26 : 11,64 = 100 : 87,78, откуда следует, что интенсивность окисления азотистых веществ ниже нормы; отношение общего количества азота мочи к количеству фосфорной кислоты выражается 13,26 : 1,00 = 100 : 14,2; отношение количества мочевой кислоты к количеству фосфорной кислоты в виде ее фосфорнодву-натровой соли выражается 0,80 : 0,68 = 1,29, что, по Цернеру, указывает на выделяемость мочевой кислоты, значительно превышающую норму; отделение общего количества фосфорной кислоты мочи к количеству фосфорной кислоты в виде ее фосфор-нодвунатровой соли как 1,88 : 0,68 = 100 : 36,2, что указывает, по Пелю, на состояние щелочности крови значительно ниже нормы.
Ввиду явных признаков бывшего мозгового поражения, больному были назначены мною с самого начала йодистый и бромистый калий, ванны в 28° R и бромистый хинин в небольших приемах. Терапия эта оказала свое воздействие и субъективно больной начал чувствовать себя лучше. Так как, однако, приступы еще продолжались и случились в период моего наблюдения — один в жаркой бане, а другой после тревоги, или, точнее говоря, после нравственных волнений, то я присоединил к предыдущей микстуре еще Adonis vernalis и Codein, а ввиду указания на мо-чекислый диатез назначено на весеннее время пить Виши. После назначения Adonis vernalis вышеуказанные приступы более не повторялись.
В приведенном наблюдении заслуживают внимания самые приступы, которые носили явный характер временной ретроактивной амнезии, так как с наступлением приступа больной, сохраняя полное сознание по отношению к своему состоянию и окружающей обстановке, тем не менее в течение приступа утрачивал совершенно воспоминание о событиях, случившихся за несколько часов перед тем и вообще не более, как за сутки по отношению к самому приступу.
При этом у больного, кроме предшествующей головной боли в задней части головы, не обнаруживалось никаких болезненных
347
ощущений и вообще больной, вполне сознававший свое состояние во время приступа, не обнаруживал никаких внешних признаков его наступления, заявляя лишь жалобы на потерю памяти недавнего прошлого. Как наступление, так и прекращение приступа не сопровождалось никакими вообще ни объективными, ни субъективными симптомами, после же приступа больной нередко, особенно при более тяжелых и продолжительных приступах, утрачивал воспоминание по отношению к самому приступу.
Последнее обстоятельство дает повод думать о присутствии в данном случае эпилептовидного состояния, которое могло развиться на почве бывшего органического заболевания мозга. В пользу этого можно было бы привести и то обстоятельство, что приступы окончательно исчезли лишь после назначения предложенной мною противопадучей смеси из Adonis vernalis с бромом и кодеином, но это обстоятельство не может быть признано существенным уже в силу того, что вышеуказанная смесь, как я убедился из своей практики, оказывает благотворное влияние и против многих других нервных заболеваний общего характера. Тем не менее, только что указанная точка зрения представляется, на мой взгляд, наиболее вероятной ввиду последующей амнезии приступа, тем более что явления ретроактивной амнезии уже описывались при падучей.
При всем том нельзя не отметить своеобразной особенности вышеописанного наблюдения, заключающейся в том, что взамен обычного, свойственного падучей симптома, характеризующегося помрачением или потерей сознания, у больного во время приступов, вместе с полной ясностью сознания и ни в чем не изменяющимся отношением его к окружающей обстановке, обнаруживается ретроактивная амнезия или забвение недавнего прошлого, стереотипно повторяющееся каждый раз вместе с развитием приступов. При этом не лишено значения, что у больного ни судорожных приступов, ни даже головокружений в течение всего болезненного состояния не обнаруживалось, что опять-таки выделяет этот случай из случаев обыкновенной падучей и заставляет признать его своеобразным эпилептоидным состоянием,, характеризующимся амнезическими приступами.
ОБ ОСОБОМ, РАЗВИВАЮЩЕМСЯ С ДЕТСТВА, НЕРВНОМ
ПОРАЖЕНИИ, ВЫРАЖАЮЩЕМСЯ ДВИГАТЕЛЬНЫМИ
РАССТРОЙСТВАМИ И СЛАБОУМИЕМ
Хотя в последний период времени наше знакомство с разнообразными наследственными формами сильно подвинулось вперед, тем не менее еще и в настоящее время мы встречаемся с типами болезней, которые не укладываются в рамки известных нам болезненных форм. Так, не далее, как в 1900 г., О. Гизе х описал особую форму наследственного нервного поражения, выражающегося дрожанием, расстройством речи и слабоумием.
В наблюдениях автора дело идет о двух больных: брата 25 лет и сестры 32 лет, у которых все прочие братья и сестры были здоровы. Больные, о которых идет речь, уже с детства обнаруживали умственную отсталось. В период же возмужалости слабоумие стало заметно усиливаться и вместе с тем начали обнаруживаться своеобразные двигательные расстройства, выражающиеся нарушением речи, которая сделалась медленной, неясной и монотонной. При этом в руках и отчасти в мышцах лица обнаружилось ясное дрожание неправильного характера, существовавшее уже при покое, при движениях же резко усиливавшееся. Вместе с тем и походка сделалась неуверенной и шаткой. Как мышечные, так и сухожильные рефлексы при исследовании обнаруживали повышение. Механическая возбудимость мышц на руках и отчасти на лице, где обнаруживалось дрожание, представлялась резко повышенной. В членах замечалась заметная ригидность. Кроме того, у сестры обнаруживался резкий парез малоберцовых мышц, у брата же паретических явлений не обнаруживалось. В общем болезнь имела медленно прогрессирующее течение. По автору, в этих случаях кора мозга несомненно должна быть поражена, равным образом автор считает вероятным и поражение мозжечка в виду существования, хотя и слабо выраженной, мозжечковой атаксии. Вместе с тем автор признает вероятным в своих случаях поражение пирамидных путей, тогда как поражение передних рогов спинного мозга он полагает возможным исключить совершенно.
1 О. G i e s e, Ueber eine neue Form hereditaren Nervenleidens Schwachsinn mit Zittern und Sprachstorung, Dtsch. Zschr. Nervenheilk., 1900, Bd. 17, Hft. 1 u. 2.
349
В том же 1900 году, к которому относится публикация О. Гизе, мне представился случай, который точно так же не подходит под описанные до сего времени наблюдения наследственных нервных поражений. Случай этот в известном отношении напоминает наблюдения, сделанные О. Гизе, но в то же время отличается от них некоторыми своеобразными особенностями, в силу чего и ввиду редкости самого заболевания он представляется интересным и заслуживает внимания. Подробное исследование этого случая было сделано по моей просьбе д-ром О. О с т а н к о в ы м. Кроме того, я сам имел возможность неоднократно наблюдать и исследовать эту больную, история которой заключается в следующем.
Больная А., 19 лет. Отец умер G? лет от скоротечной чахотки после инфлуешш; он был очень нервный. Мать страдает грыжей. Братья отца были люди с большими странностями. Тетка ло отиу страдала душевным расстройством и умерла в больнице на Удельной. Отец был женат два раза. Наша больная ■— дочь от второго брака. От первого брака отец имел троих детей, из которых одна дочь страдала базедовой болезнью, один сын пил запоем, спился и умер от чахотки. От второго брака, кроме пашей больной, есть еще один сын который отличается нервностью. Со стороны матери дед и дядя были недостаточно развитые, дядя, кроме того, страдал падучей болезнью.
Родилась больная недоношенным ребенком (на 8-м месяце) и в асфиксии, вследствие того, что мать не поберегла себя во время беременности от тяжелых физических усилий и имела повод подвергаться волнениям. После рождения девочка страдала еще кровотечениями из пуповины в течение месяца, вследствие плохой ее перевязки. Зубы прорезались в срок, ходить начала больная около году и четырех месяцев; ходила однако не твердо и часто падала. Хорошей, правильной походкой она никогда не обладала. До 4 лет больная ничего не говорила. Затем после перенесенной тяжелой формы скарлатины больная стала бойко говорить, а 6 лет самоучкой выучилась читать; окружающие ее родные в это время замечали у нее какое-то ненормальное нервное возбуждение.
До 9 лет больная перенесла серию заболеваний, сильно ее ослабивших, как-то: корь, коклюш, ветреную оспу, а 9 лет, по словам матери,, перенесла «воспаление мозга», после чего она уже не могла учиться, забыла все, что учила раньше, и затем так и осталась недоразвитым физически и психически ребенком. 11 лет показались менструации. В этом же возрасте у больной замечена была ненормальная наклонность ко сну и обнаруживалась раздражительная слабость. До 1899 г. мать ее считала все-таки психически здоровой, хотя и отсталой в умственном отношении. В 1899 г. в сентябре больная заметно стала хуже ходить, стала чаще падать и однажды вывихнула себе ногу, лечение которой взяло шесть недель. Со времени этого вывиха больная стала очень раздражительной, часто стала говорить о своей непригодности к жизни, жаловалась на предсердеч-ную тоску и высказывала желание поскорее умереть.
Зимой в 1899 г. больная успокоилась и даже мечтала о замужестве. Осенью 1900 г. она стала опять раздражительной, стала по целым ночам плакать, грозилась покончить с собой, но попыток к самоубийству нк разу не обнаруживала. Ничем больная не занимается, кроме игры в карты. Часто больная сердится, а по временам бывает значительно возбуждена.
Status praesens. Окружность черепа 54 см. Ушные раковины отстоят ст черепа значительно, ушные сережки выражены слабо, левая ушная раковина несколько больше, чем правая. Зрачки неравномерны, левый шире правого, на свет зрачки реагируют хорошо. Язык тоньше нормального, при высовывании замечается резкое дрожание. При осмот-
350
ре больной должно отметить большое отложение жира в подкожной клетчатке; груди отвисшие, развитие грудей обращает на себя внимание в виду их чрезвычайно больших размеров. Руки в кистях производят впечатление недоразвитых, в пальцах рук наблюдается уже при покое мелкое резко выраженное дрожание, которое еще более усиливается при движении; писать больная может с трудом, при чем почерк ее неровный, дрожащий (см. рис. 1). Кисти рук резко цианотичны, покрыты клейким потом и холодны на ощупь. Мелкие движения пальцев медленного темпа; скоро больная выполнять их не может, вследствие некоторой ригидности мышц. Мышечная сила правой руки 48 фунтов, левой 50 фунтов. Все движения в предплечьях и плечах вполне возможны. Предплечья, как и кисти, развиты слабее, чем плечи. Походка больной колеблющаяся и спастически-иаретическая; больная заявляет, что при ходьбе йоги ее не сгибаются; при этом туловище ее держится не прямо, а несколько согнувшись. Вместе с тем при ходьбе больная заметно отклоняется в правую сторону и вообще походка больной не твердая и несколько колеблющаяся.
Рис. 1.
Объем стоп: правой 25 см, левой 24 см; объем правой голени 38 см", левой 40 см, объем правого бедра 5G см, левого 58 см. Стопы и голени развиты слабее, бедра же чрезвычайно толсты. Стопы холодны на ощупь к-кожа имеет красный оттенок. Мышечная сила ног ослаблена, особенно это. выражено в сгибателях стопы и разгибателях голени. Возбудимость мышц. без резких изменений.
Исследование болевой чувствительности не обнаруживает никаких расстройств; тоже следует сказать о тактильной н температурной чувствительности. Позвоночник в шейной и плечевой части изогнут кзади более нормального.
Кости черепа чувствительны к постукиванию, особенно с левой стороны. Позвоночник к постукиванию не болезнен. Подошвенный рефлекс выражен хорошо, ягодичный не получается, брюшной и подчревный также. Сухожильные рефлексы верхних конечностей ослаблены.
Рефлексы с ахиллова сухожилия получаются; коленные рефлексы также получаются, но слабо. Клонуса коленной чашки и клонуса стоп нет.
Со стороны внутренних органов следует отметить слабые тоны серд ца и легко сжимаемый пульс (70 в 1'). Со стороны органов чувств больная заявляет жалобы на ослабление зрения. Со стороны мочеотделения больная жалуется на бывающие у нее по временам частые позывы. Отправления кишечника вялы.
Со стороны психической у больной замечается резкая отсталость в развитии, она не может сказать какой год, не может прочесть многозначных чисел и пр. Память умеренная. Настроение духа большею частью подавленное. Никаких бредовых идей больная не высказывает. Речь больной не вполне ясная, несколько лепечущая. Восприятие замедленное.
Свое настроение она объясняет тем, что над ней смеются и обижают ее, поэтому она и чувствует тоску, раньше же помышляла о самоубийстве. Беспомощность ее состояния всегда ее сильно удручала. По временам больная вспоминает все печальное из прошлого, изображает в жалком виде себя, говорит, что ее били, не любили и обижали. Под влиянием таких мыслей у нее наблюдаются приступы неудержимых рыданий. Чувственная реакция у больной вообще сильно повышена. Обманов чувств не наблюдается.
351
Рассматриваемое заболевание по внешности, ввиду явлений мозжечковой атаксии и расстройства речи при отсутствии изменений чувствительности и при медленно развивающемся с раннего возраста страданий, может напоминать в известной мере наследственную атаксию Фридриха, но оно отличается от нее отсутствием явлений мышечной атаксии, хореатических явлений и нистагма, сохранностью коленных сухожильных рефлексов, дрожанием членов, некоторой ригидностью и явлениями слабоумия, не говоря уже о наблюдавшемся в нашем случае недоразвитии периферических частей конечностей.
Что касается наблюдений, сделанных О. Гизе, то есть достаточно оснований признать, что в нашем случае дело идет о подоб ном же своеобразном заболевании. Но наш случай выделяется от случаев О. Гизе, тем, что сухожильные рефлексы не представлялись повышенными, равным образом и повышенной мышечной возбудимости в нашем случае не наблюдалось. С другой стороны наш случай выделяется от наблюдений О. Гизе ясным недоразвитием периферических отделов конечностей, отсутствием местного паралича, как было бы по крайней мере у больной сестры О. Гизе, и более или менее резкими явлениями застоев в конечностях. Нельзя не обратить внимания также на то обстоятельство, что в нашем случае, кроме крайне неблагоприятной наследственности, имелась совершенно особенная этиология заболевания в виде преждевременных родов и целого ряда других, бывших в детстве больной моментов, окончательное значение которых в отношении развития болезненного состояния должно предоставить будущему.
Хотя болезненное состояние в нашем случае заметно стало обнаруживаться около 9-летнего возраста, со времени появления тяжелых мозговых симптомов, но дегенеративные признаки и недоразвитие конечностей у больной ясно свидетельствуют, что первоначальные задатки болезни должны быть даны уже неблагоприятными условиями наследственности, тем более что походка больной уже с самого начала представлялась неправильной. Тем не менее должно иметь в виду, что развившееся в 9-летнем возрасте мозговое поражение у нашей больной впервые дало существенный толчок к развитию ее болезненного состояния, с тех пор медленно, но постоянно прогрессирующего. Имея в виду наличность у больной признаков слабоумия, резкого, дрожания конечностей и своеобразного расстройства походки с характером мозжечковой атаксии, должно признать, что дело идет в данном случае о поражении мозговой коры и мозжечка, причем, как надо думать, подвергаются вторичному перерождению как пирамидные, так и мозжечковые нисходящие пучки.
О НАВЯЗЧИВЫХ КИШЕЧНЫХ И ПУЗЫРНЫХ КРИЗАХ И О ЗНАЧЕНИИ ПРИ НИХ ЛЕЧЕНИЯ ВНУШЕНИЕМ
В заседании от 28 октября 1904 г. я сообщил научному собранию клиники душевных и нервных болезней свои наблюдения относительно пузырных и кишечных кризов навязчивого характера.
В этом сообщении мной были указаны и те немногие литературные данные, которые имелись по занимающему нас вопросу, начиная с Оппенгейма1 и затем указания Нотнагеля, Боаса2, Паризера3, Пика4, цитированные Оппенгеймом в статье, относящейся к этому предмету и помещенной в Медицинском обозрении за июнь, т. LV5.
В настоящем сообщении я остановлюсь еще раз на своеобразных расстройствах кишечника, которые, на мой взгляд, правильнее всего могут быть названы навязчивыми «кишечными расстройствами, или кризами».
Дело идет в этом случае о настоящих расстройствах кишечника с частыми позывами и даже поносом в такое именно время, когда расстройство кишечника по обстоятельствам дела является крайне неуместным. Благодаря этому, оно нарушает в такой мере жизненные условия, что держит больных под постоянным страхом за возможное развитие кишечного расстройства в период времени, когда оно является особенно неудобным.
Но приведем здесь самые наблюдения, относящиеся к занимающему нас предмету и представляющие собою несколько типичных случаев, бывших под моим наблюдением за последнее время.
Больная У., 32 лет, замужняя. Более 10 лет начала страдать навязчивыми кишечными кризами. Она уже с юности имела нервные кишечные
1 О р р е η h e i га, Handbuch, S. 796.
2 Boas, Diagnostik u. Therapie d. Darmkrankheiten, 1899.
3 P a r i s e r, Chronische nervose Diarrhoen u. ihre Behandlung, D. med. Zeitung, Berlin, 1900.
4 Pick. Ober eine psychisch bedingte Slorung d. Defektion, Wiener klin. Wschr., 1901, № 20.
вОппенгейм, проф., О психическом характере некоторых расстройств актов мочеиспускания и испражнения, Медицинское обозрение, июнь 1901.
23 Б. Л\. Бехтерев
353
расстройства. Во время ее пребывания в Риме у ней началось случайно расстройство кьшечнлка, вследствие которого она должна была выходить из-за табльдота. Это ее сильно стесняло, и она немало волновалась. Но чем больше она волновалась, тем больше ее беспокоило расстройство кишечника и тем чаще являлись позывы.
С этого времени и начала обнаруживаться боязнь кишечного действия при всевозможных случаях, когда такое действие может оказаться неуместным.
Больная сама о начале своей болезни пишет следующим образом: «Я родилась недоноском и с раннего детства была уже нервною, всегда плохо спала и была очень впечатлительной.
Когда мне было около 19 лет, вследствие болезни отца t a b e s' ом, дома была постоянно обстановка больницы. На меня это наводило смутный страх и вызывало безнадежность. 13г/„ лет меня отдали в закрытое заведение, и я стала усиленно учиться; через 2 месяца я сформировалась и вскоре после этого заболела тяжелой скарлатиной. После нее стала постоянно худеть, очень ослабела, появились невралгии, головные боли. На следующую осень перенесла сильную корь.
С тех пор каждый менструальный период сделался настоящей болезнью, гричем первые дни всегда делалось сильное кишечное расстройство. Во время экзаменов являлось такое же расстройство, которое тогда объясняли простудой, но я теперь убеждена, что это было нервное состояние.
По окончании гимназии меня стали «вывозить», и каждый переход от тягостной монотонной и одинокой жизни в деревне к слишком разнообразной жизни в П-ге вызывал опять мой своеобразный «катарр кишок». Но тогда это скоро проходило, особенно после солевых согревающих компрессов.
19 лет меня повезли за границу. Едва переехав в Рим из Флоренции, я почти сейчас же заболела желудком в прежнем роде, но на этот раз болезнь приняла особенную форму.
У себя в комнате, во время катанья, гулянья я была совсем здорова, по стоило сойти в ресторан к табльдот или пойти в театр, как меня начинали мучить схватки. И чем я больше этим стеснялась, чем неудобнее было выйти, тем эти позывы были сильнее и чаще. Ни лекарства, ни красное вино — ничто не помогало.
И с тех пор более 10 лет эта болезнь не поддается никакому лечению.
Боясь этих схваток, я одно время просто морила себя голодом и ужасно ослабела. Тогда решили, что у меня римская малярия; замучили мышьяком (уколами), железом, посылали в Аббацию.
В это время я вышла замуж, и вопреки мнению докторов мне стало еще хуже.
Одно время, не то от жизни на юге в полном спокойствии, не то от теплых морских ванн мне стало лучше, но скоро после этого наступила беременность, которая вызвала ужасное нервное состояние, стремление к самоубийству и потребность причинять себе и окружающим боль и вред.
Таким образом, беременность и роды меня только истощили, и мое состояние опять ухудшилось, хотя проф. Р. и др. говорили, что у меня просто истерия, вызванная неправильным положением и плохим развитием женских органов, и что «материнство меня вполне исцели т», но, считая его маловероятным, меня убеждали согласиться на операцию.
С тех пор я и лечилась, и пробовала увлечься делом, и утомляла cc/:i физическим трудом или только лежала и ела (у Лейдена), но ничто ье-помогало.
При вполне спокойном состоянии желудок действует правильно, как часы, скорее всегда крепок, но стоит начать собираться, особенно спешно, в театр, в церковь, на вокзал, — тотчас является позыв за позывом и желудок действует 5—6 и более раз.
В обществе же и от музыки большею частью является чувство расписания газами и невозможность их сдержать. Иногда, хотя и редко, они
3>!
отходят отрыжкой. Слишком сильное усиление сдержаться вызывало тогда нервные припадки; дрожь и подергивание всего тела или рыдания. Иногда крепкое вино или что-нибудь очень кислое помогает пересилить позывы, но с годами это все становится труднее.
Дед мой по отцу — мечтательная, артистическая натура. После одной неудачи поселился в деревне, стал тосковать и пить. В этом уже периоде он женился и кончил жизнь самоубийством.
Отец очень даровитый (талант к живописи), но рано повел «широкую» жизнь и в 25—26 лет уже страдал ипохондрией, а порой галлюцинациями.
Женился 35 лет уже больной tabes' ом. Мать вечно страдала мигренями и нервами, потом подагрой.
Сестра матери не вполне нормальная.
В период моего наблюдения у больной появляются позывы на низ и расстройство кишечника, как только куда-нибудь ей приходится ехать, например, в театр, в церковь, к знакомым и т. п., а также и в том случае, когда она ждет к себе знакомых. Уже при одевании ей приходится постоянно бегать в клозет, что случается от 5 до 10 раз, прежде чем она выедет. С ней случается иногда, что, приехав к своим знакомым и позвонив к ним, она вынуждена возвращаться домой для того,чтобы еще раз итти в клозет. По словам больной, однажды в театре она должна была спешно выбежать из кресел и заплатить большие деньги, чтобы мгновенно была сломана запертая дверь в клозете.
В другой раз во время пожара, когда нужно было бы спасать себя и имущество, она должна была сидеть в клозете. Чем более важно для больной быть где-нибудь, тем и позывы мучительнее.
Если больная при этом удерживается, то у ней начинается вздутие живота, очень мучительное и тягостное, с громким ворчаньем,-которое ее немало стесняет в гостях. Вообще она делает иногда невероятные усилия, чтобы пересилить себя, но все же преодолеть позывы часто не в состоянии никакими средствами. Заслуживает внимания, что у больной даже пение, особенно церковное, а также музыка, преимущественно высокие ноты, возбуждая нервное состояние, вызывают вместе с тем и вздутие кишечника. В обыкновенное же время действие желудка бывает нормальное.
Самые разнообразные меры, применяемые против болезненного состояния, оказались безуспешными. Некоторую пользу больная видела лишь от капель опия в период расстройства кишечника. Но на развитие позывов в гостях или в общественных местах это почти не имело влияния. Скорее в этом смысле оказывали некоторое влияние приемы спиртных напитков. Но вино ей помогает лишь на час или два, а затем она чувствовала от него лишь усталость и слабость
В общем же надо отметить, что больная, лечась в течение (О лет почти беспрерывно, не получила заметного облегчения. Скорее ее состояние постепенно ухудшилось.
Объективно у больной обнаруживается дрожание век при их закрытии, некоторое повышение сухожильных рефлексов
23*
355
и небольшое искривление матки. В остальном никаких расстройств. Исследование испражнений на яйца глистов дало отрицательные результаты.
Больной П., молодой человек, происходит от истеричной матери и нервного отца. В прежнее время имел, по определению проф. Пастернацкого, катарр кишечника, от которого излечился. Со временем у него стали появляться схватки в кишечнике и позывы к жидкому испражнению при том или другом волнении. Однажды, готовясь в академию, он подумал, что это расстройство не даст возможности ему поступить. При одной этой мысли уже появляются схватки в животе, позыв к пище, за которым следует жидкое испражнение. Затем он стал опасаться, что подобное же с ним может случаться при других условиях, особенно при неподходящей обстановке. И действительно, эта мысль стала его беспокоить при всевозможных необычных условиях. Так, когда он бывает в гостях, то у него уже задолго До того являются мысли со страхом за возможность неприятного положения. В гостях его сильно беспокоят газы, которые бродят в кишечнике, и его начинает все сильнее и сильнее тяготить мысль о газах, хотя больной все же с трудом может удерживаться. С объективной стороны ничего заслуживающего внимания.
Больная 23 лет, девица, имеет нервных отца и мать, сама обнаруживает общую нервность уже с 7 лет и с того же времени заикание; в детстве имела истерические конвульсии. Вместе с тем она имела всегда склонность к поносам. Лет 10—15, как начали обнаруживаться частые позывы и наклонность к поносам при всяком волнении. Вследствие этого развилась настоящая боязнь кишечных расстройств в разных случаях, когда это признается неудобным. В театрах всякое вообще волнение, будет ли это радость или страх, вызывает тотчас же позывы, и она должна выходить для пользования клозетом. То же самое случается и в гостях. При обычных условиях, когда больная не испытывает волнения, у нее не бывает и частых позывов. Если она удерживается от позывов во время волнения, то у нее наступает вздутие живота «и такое ворчание, которое слышно в другой комнате».
При объективном исследовании, кроме небольшого повышения коленных сухожильных рефлексов, не открывается каких-либо особых явлений со стороны функций нервной системы.
Больной Б., 36 лет, женат, управляющий нефтепромышленной фирмы, происходит от нервной матери, страдающей ревматизмом и ожирением сердца. Отец умер от болезни сердца. Другие родственники в нервном и душевном отношении здоровы. Больной развивался правильно. Сифилиса не имел, имел лишь умеренный онанизм. Около 10—14 лет жалуется на сильную бессонницу, причем поводом к пробуждению обыкновенно является позыв к мочеиспусканию. Издавна больной страдает плохим пищеварением, отрыжками после приема пищи и поносами, являющимися на смену запоров. Еще раньше появления бессонницы, лет 15—16 тому назад, без всякой видимой причины у больного начались расстройства кишечника и склонность к частому мочеиспусканию, особенно когда он озябнет. Летами же обыкновенно больной чувствовал себя хорошо. Затем начались созывы к мочеиспусканию и испражнению при всяком волнении. Особенно же мучают больного позывы при такой обстановке, когда выполнение самого акта становится неудобным, например в зрительном зале театра, в незнакомом обществе и т. п. По заявлению больного, при голоде эти позывы вообще его беспокоят сильнее, нежели при сытом состоянии. Больной уже вперед мучается мыслями о возможном появлении позывов в обществе и, когда они действительно наступают, то при неудовлетворении потребности дело доходит до похолодания конечностей и дрожания рук. Из объективных симптомов можно отметить некоторое повышение коленных рефлексов; пульс 78 в 1 минуту; умеренный дермографизм.
356
В остальном ничего патологического ни со стороны чувствительности, ни со стороны органов чувств, ни со стороны движения, ни со стороны recti и мочеполовых органов, кроме вышеуказанного расстройства.
Я не увеличиваю числа наблюдений только потому, что не все случаи, относящиеся к амбулаторным больным, записаны так подробно, как предыдущие.
Если подвести теперь итог вышеприведенным и другим наблюдениям относительно того своеобразного расстройства, которое мы можем назвать «навязчивыми кишечными кризами», то мы убедимся, что почти во всех случаях дело идет о больных, происходящих из невропатической или психопатической семьи.
Второе обстоятельство, которое в этом случае, повидимому, не лишено известного значения, заключается в том, что все или почти все больные издавна страдают неправильностями в отправлениях кишечного канала, основанными, по крайней мере в некоторых случаях, на бывших ранее поражениях желудочно-кишечного канала, особенно толстых кишок.
Как выражение установившейся связи между нервным состоянием и расстройствами кишечника у больных обнаруживаются кишечные расстройства при каждом душевном волнении. Этим волнением, возбуждаюшим кишечные расстройства, собственно и дается почва для частых позывов к испражнению, которые развиваются у больных при тех условиях, когда выполнение отправления кишечника представляется по внешним обстоятельствам неудобным или даже неисполнимым. Боязнь кишечных расстройств, неправильно называемая некоторыми «клозето-боязнью», обыкновенно является на почве вышеуказанных состояний, когда одна мысль о невозможности выйти из затруднительного положения при появлении позывов сопровождается волнением и приводит к появлению частых позывов. Эта мысль, как волнующая, становится навязчивой и начинает беспокоить больных уже вперед при одном предвидении неудобного положения в обществе от возможных позывов, когда является необходимость быть среди общества. Что касается «навязчивых пузырных кризов», то, как показывают наблюдения, они представляют собою расстройство, которое лишь в отдельных случаях наблюдается совместно с «навязчивыми кишечными кризами», чаще же представляют, как я убедился из других бывших в моем распоряжении наблюдений, самостоятельное нервное расстройство, аналогичное по своим особенностям и внешним проявлениям «навязчивым кишечным кризам». Часто и здесь дело шло о больных, которые, вследствие тех или других условий, приведших к раздражению шейки мочеиспускательного канала, страдали частыми позывами на мочу, и это обстоятельство, по-видимому, служило первоначальным поводом к развитию волнения в случаях появления позывов во время пребывания в обществе и к развитию навязчивой идеи и страха за возможность появления позывов на мочу в гостях или в собрании.
357
Оба вышеуказанные расстройства, хотя и наблюдаются у невропатических субъектов и могут развиваться у настоящих неврастеников, но прямой зависимости от неврастении не имеют, так как могут наблюдаться и при отсутствии ясно выраженной неврастении.
Что касается лечения, то в моих случаях оказывало пользу и давало благоприятный результат комбинированное лечение психотерапией и физическими методами. Из последних больным назначались успокаивающие нервную систему средства вместе с применением опиатов. Удачной комбинацией для этой цели оказалось применение бромов с кодеином в достаточных приемах и эфирная валериана с каплями строфанта или других сердечных, действующих успокоительно на приступы сердцебиения, развивавшегося при волнениях.
Наряду с этим небесполезными как успокаивающие нервную систему оказались тепловатые ванны.
Но существенную часть лечения в моих случаях составляла психотерапия в виде систематически проводимых внушений. Значение психотерапии может быть прослежено, между прочим, на первом наиболее тяжелом случае, окончившемся, благодаря внушению, полным выздоровлением.
Когда эта больная обратилась ко мне за советом, она просила лишь облегчить ее состояние в какой-либо мере, не надеясь ничуть на полное выздоровление. Ей было назначено по обыкновению комбинированное физическое и психическое лечение. С этой целью для успокоения нервной системы ей предложено принимать смесь из брома с кодеином, валерианом и строфантом, тепловатые ванны и вместе с тем предложено лечиться внушением. К последнему больная обратилась лишь несколько времени спустя, когда убедилась, что вышеуказанная физическая успокоительная терапия, успокоив нервы и ослабив временами появлявшиеся кишечные расстройства, не дала существенных результатов по отношению к основному поражению, выражающемуся навязчивыми кризами. Для устранения последних производились внушения, которые могли быть осуществлены в состоянии легкого сна. После первого же сеанса больная почувствовала улучшение, так как после него могла быть в театре и в гостях, забывая совершенно о своем недуге.
После ряда внушений состояние здоровья настолько улучшилось, что больная могла совершенно свободно быть в гостях или в собраниях до ночи без всяких намеков на прежние кишечные кризы; лишь перед отъездом в гости или в собрание она должна была раз или два для своего успокоения побывать в клозете, что ее нисколько не тяготило.
При дальнейших внушениях и этот остаток болезни исчез, и больная признала себя выздоровевшей от своего недуга.
О РАССТРОЙСТВАХ В СФЕРЕ ЧУВСТВЕННОГО ВОСПРИЯТИЯ У ДУШЕВНОБОЛЬНЫХ
Психические расстройства, приводящие к понижению или обострению чувственного восприятия или к возникновению самостоятельных ощущений элементарного характера в виде болей и других каких-либо ненормальных ощущений, без сомнения, не остаются без значения в общем симптомокомплёксе душевных расстройств. Но во многих отношениях они еще не могут считаться хорошо изученными, вследствие чего я и полагаю возможным остановиться на них в настоящей работе1.
Рассматриваемые явления в известном смысле напоминают собою физические изменения чувствительности или обыкновенные анестезии, гиперестезии и невралгии, но в отношении своего патогенеза отличаются от них тем, что они вызываются у душевнобольных не какими-либо нарушениями в Проводящей деятельности нервной системы, а обусловливаются психическим состоянием больных, являются, таким образом, психически обусловленными расстройствами ощущения и потому в отличие от первых могут быть названы психическими анестезиями, психическими гиперестезиями и психическими алгиями или короче психоанестезиями, психогиперестезиями и психоалгиями.
Психоанестезии выражаются более или менее ясным ослаблением чувственного восприятия и могут наблюдаться как в сфере общей чувствительности, так и в сфере органов чувств. Они обусловливаются либо общим понижением сознания у душевнобольных, либо отвлечением их внимания болезненными явлениями. Прототипом первых в нормальном состоянии является то ослабление чувственного восприятия, которое наблюдается у здоровых людей во время дремоты или сна, прототипом второго — то ослабление чувственного восприятия, которое мы наблюдаем у всех здоровых лиц при отвлечении внимания. Известно, что при серьезном умственном труде заглушаются даже такие тягостные ощущения, как зубные боли; равным образом в пылу сражения или общей свалки люди не замечают полученной ими раны.
1 Краткие указания на эти расстройства мы встречаем у Эббипгауза, Крафт-Эбинга и др.
359
Совершенно подобные же явления наблюдаются нере.ш-о ь у душевнобольных. Таким образом, при сноподобных психических состояниях, которые мы наблюдаем, например, при amentia Meynerii и при некоторых эпилептических формах помеша-гельова, чувственное восприятие оказывается ясно пониженным, благодаря чему внешние раздражения кожной поверхности и впечатления, действующие на органы чувств, не воспринимаются с той отчетливостью, как это бывает в нормальном состоянии.
В этом нетрудно убедиться при непосредственном наблюдении больных и при испытании их кожной чувствительности; наконец, и сами больные при улучшении их состояния говорят о более или менее общем притуплении сферы чувственного восприятия.
С другой стороны, когда больные заняты своим бредом, когда их внимание отвлечено теми или другими идеями, то естественно, что под влиянием этого чувственное восприятие ослабевает и обнаруживается более или менее ясно выраженная психоанестезия. Наивысшей степени это состояние достигает во время экстаза при религиозном возбуждении, но его можно, наблюдать и при таких состояниях, когда внимание больных отвлечено обильными слуховыми, зрительными или иными галлюцинациями.
В некоторых случаях эти психоанестезии приобретают систематический характер, сбнаруживаясь в отношении одного какого-либо рода ощущений.
Так, иногда мы имеем психоанестезии в сфере общего чувства, наблюдаемые, между прочим, нередко при истерии, хотя они встречаются и при других, особенно при органических психозах. Больные такого рода иногда сами жалуются на пустоту в голове, на то, что они не чувствуют своей головы, своего тела, своих внутренностей и т. п., несмотря на то, что специальное исследование кожной чувствительности и чувствительности глубоких частей тела не обнаруживает в этом отношении никаких существенны х изменений.
Само собой разумеется, что такого рода психоанестезии в известных случаях приводят к развитию своеобразного бреда об отсутствии внутренностей, о наступлении смерти и т. п.
Нижеприводимый случай кататонии представляет один из классических примеров резко выраженной психической анестезии в сфере общего чувства.
Больная — дочь купца, девица 20 лет, происходит из здоровой семьи. Ни тяжелых нервных, ни душевных заболеваний среди ближайших родных нет. Отец 60-летний здоровый старик. Мать 50 лет; иногда страдает сильными сердцебиениями. Дед со стороны матери 20-летний здоровый старик. Бабка со стороны матери, 88 лет, здоровая женщина. Дед и бабка со стороны отца умерли в преклонном возрасте от неизвестных причин. Больная седьмой ребенок, самая младшая. Имеет двух братьев вполне здоровых и четырех сестер, из которых одна, самая
зео
старшая, очень нервная, впечатлительная особа. Дети братьев и сестер вполне здоровы.
До первого года жизни больная была вполне здорова. На втором году у больной сделался язвенный процесс на голени правой ноги. Нога болела около 2 лет. Сделали две операции. Вследствие этой болезни больная сею жизнь хромает. Хромота заставляла больную избегать общества, так как она боялась показаться смешной. Она была всегда почти грустна и раздражительна. До 16 лет ничем другим не хворала. С 17-летнего возраста стала страдать малокровием и сильными сердцебиениями. Года 2 тому назад стала страдать глазами: сначала заболел притом правый глаз, затем левый. Глазная болезнь тянулась хронически с частыми обострениями, сопровождаясь сильными болями. Последнее обострение глазной болезни было весной 1901 г. Больная стала высказывать опасение ослепнуть. В мае месяце настоящего года приехала из Киева в С.-Петербург для совета с глазными врачами. Около этого же времени обострилось и ее нервное заболевание. Обнаружилась сильная слабость, появилась частая рвота, подкатывание шара к горлу. Лечилась подкожными впрыскиваниями мышьяка. В начале июня появилась упорная бессонница, больная стала тосковать, беспокоиться, высказывать идеи, что она не чувствует своей головы, у нее нет рта, она не чувствует, сыта ли она. За самое последнее время болезнь ухудшилась. У больной появились мысли, что она уже умерла.
Больная роста ниже среднего, плохого питания. Наружные покровы лица землистобледного цвета. Видимые слизистые оболочки слегка анемичны за исключением конъюнктивы обоих глаз, которая представляет резкие явления гиперемии. Мышцы развиты умеренно. Костная система развита правильно, за исключением правой большой берцовой кости, которая представляется неровной и несколько болезненной при ощупывании. На наружной поверхности правой голени линеарный рубец от бывшей секвест-ротомии. Лицо без асимметрии. Левый зрачок уже вследствие задних синехий. Следы бывшего ирита в обоих глазах. Реакция зрачков на свет в правом глазу хорошая, в левом несколько ослаблена. Подвижность глазных яблок нормальная. Коленные рефлексы слегка повышены, причем левый несколько сильнее правого. Сухожильные рефлексы на верхних конечностях выражены ясно, равномерны с обеих сторон. Глоточный рефлекс и справа, и слева сохранен. Болевая чувствительность без изменений. Мышечная сила ослаблена. Походка с прихрамыванием иа правую ногу. Тоны сердца чисты. Пульс 76 ударов в одну минуту, правильный, умеренного напряжения. Живот втянут, стенки его вялы. Упорный запор. В моче ни белка, ни сахара нет.
На лице больной написано тоскливое беспокойство; она беспрестанно как бы с испугом озирается по сторонам. Все время сидит на постели и держит свою голову левой рукой за подбородок. Плаксивым голосом умоляет врача помочь ей и непременно тотчас же, просит сказать ей, излечима ли ее болезнь, умоляет спасти ее, так как ей грозит гибель. Всецело поглощена своими идеями, плохо воспринимает вопросы, не относящиеся к сфере ее бреда; она на них или ие отвечает вовсе, или, начав отвечать, прерывает просьбами помочь, спасти и т. д. Больная утверждает, что ей кажется, что у нее нет головы, нет рук, нет ног, что в середине ее нет ничего. По временам она говорит, что голова у нее отделена от сердца, «заверчена болезнью назад» и совершенно очумела. В ней нет мыслей, нет памяти, нет желаний, радостей, горя и пр. Больная говорит, что она не может ходить, не может лежать, не может сидеть, в то же время по просьбе врача тотчас встает, ходит и т. д. Мучительно сознает невозможность что бы то яи было сделать, что-нибудь пожелать. Утверждает, что не может есть, что не чувствует голода, не чувствует пищи. При побуждении же к еде, при кормлении кушает достаточно. Уверяет, что не спит воЕсе. Ей кажется, что она сильно похудела, что остались только кости, что и костей она уже не чувствует, что с каждым днем ее состояние ухудшается, что она неизлечима. Думает, что настоящая болезнь ее произошла оттого, что ее ле-
361
чили от глазной болезни ртутными втираниями. Больная постоянно держит себя за голоьу, чтобы убедиться, что голова у нее есть, но этого мало, и больной все-таки кажется, что головы у нее нет, что она ее не чувствует. Больная утверждает, что из всего ее организма только и действует ее язык. Она думает «с помощью языка», боится «с помощью языка», желает, понимает «языком» и т. д. Когда же она перестает говорить, то она перестает быть живой, становится, «как покойница».
При испытании и расспросах выясняется, что чувствительность (осязательная, болевая, мышечная и пр.) у больной вполне сохранена. Она сама заявляет, что чувствует свое тело и голову, когда прикасается к ним своей рукой, и тем не менее ей кажется, что ни головы, ни туловища у нее нет, что она их не чувствует, хотя и убеждена, что они все же существуют. Давление на внутренности в животе также больная ощущает, но тем не менее сама она их не чувствует. С руками то же самое: больной кажется, что их нет, тогда как на самом деле и кожная, и глубокая чувствительность их (мышц, костей, связок) вполне сохранена, и она убеждена, что руки у нее есть, она думает, однако, что они сделались у нее легкими и потому она не испытывает их тяжесть, производя на глазах всех махательные движения рук.
Относительно приема пищи больная также утверждает, что сна не чувствует, что она ест, не чувствует, что насыщается и т. п. Она не чувствует горя и радости, когда же обращают ее внимание на то, что она охает или стонет, то она говорит, что это «языком только».
Что касается постоянного держания рукой своей головы за подбородок, то больная говорит, что, хотя это и не дает ей ощущения головы, но оиа по крайней мере знает, где находится ее голова. Со временем у больной развилось неподвижное, периодами даже каталептоидное состояние, с мыслями о том, что оиа умерла, иногда сопровождаемое полной немотой с отказом от пищи. Спустя несколько недель больная снова выходит из своего неподвижного состояния, обнаруживая постепенное улучшение своего состояния. Таким образом, в данную пору развития болезни мы, несомненно, имеем у больной анестезию в сфере общего чувства, но не физическую,как показывает соответствующее исследование^ психическую.
Само собой разумеется, что такого рода анестезия может приводить, как это и наблюдается у нашей больной, к бредовым идеям соответственного характера, т. е. к бреду об отсутствии внутренностей, к бреду о наступлении смерти и т. п. Интересно отметить, что, наряду с психической анестезией, у нашей больной имелась и психическая анестезия душевного чувства. Несмотря на то, что больная, видимо, страдает, выражая свои страдания охами, стонами, мимикой и просьбами о помощи, она тем не менее утверждает, что она не чувствует и не способна чувствовать ни горя, ни радости, а если она и заявляет просьбы о помощи, то это только языком. Как известно, нечто подобное наблюдается иногда и при тяжелых формах меланхолии.
Что касается противоположного состояния — психической гиперестезии, то в противоположность психическим анестезиям она обусловливается или общим обострением всех психических процессов и повышением их яркости, или же специальным сосредоточением внимания на тех или других впечатлениях. Такие гиперестезии наблюдаются в сфере кожной чувствительности и органов чувств при маниакальном возбуждении, а равно и при всех состояниях психического возбуждения, обусловливаясь здесь главным образом обострением психических процессов.
362
При первичном сумасшествии, благодаря систематическому бреду и галлюцинациям, обыкновенно наблюдаются психические гиперестезии под влиянием сосредоточенного внимания на определенном круге идей, являющихся, следовательно, систематическими.
Примером таких систематических психических гиперестезии может служить один бывший под нашим наблюдением офицер, получивший контузию в прошлую русско-турецкую войну и с тех пор страдавший травматическим неврозом с психическим расстройством. Он не переносил слова «кровь» и обнаруживал резкую гиперестезию по отношению ко всему красному. При слове «кровь» и при виде чего-либо красного он приходил в страшное волнение и с ним развивались настоящие судороги. Психические гиперестезии в сфере общего чувства также нередки у душевнобольных и выражаются чувством общей разбитости, обострением чувства голода и чувства сытости и т. п. Но особенное значение получают психические гиперестезии, как известно, при ипохондрических состояниях и при ипохондрическом сумасшествии, при которых ощущения от внутренних органов, в нормальном состоянии неясно сознаваемые, не только хорошо сознаются больными, но и становятся тягостными для них.
Нельзя, конечно, отрицать, что при этих состояниях нередко причиной гиперестезии внутренних органов служат те или другие поражения этих органов (желудочно-кишечный катарр, застои в области воротной вены и т. п.) и, следовательно, здесь может итти дело в известной мере о физической гиперестезии. Но, несомненно, также, что, наряду с такой физической или проводниковой гиперестезией, при этих состояниях наблюдается и психическая гиперестезия в сфере общего чувства, так как достаточно бывает такого рода больным направить внимание на свои внутренние органы, чтобы уже испытывать в них те или другие ощущения. Развитие бреда на основании этих психических гиперестезии в виде гниения или разложения внутренностей — факт слишком общеизвестный, чтобы на нем останавливаться.
Возникает, конечно, существенный вопрос, как отличать психические анестезии и гиперестезии от анестезий и гиперестезии физических или проводниковых. Насколько позволяет мне судить в этом отношении мой собственный опыт, важными отличительными признаками первых от последних служит отсутствие каких-либо указаний на нарушение проводимости нервной ткани, обусловливающее проводниковую или физическую анестезию или гиперестезию, с другой стороны, их общий или систематический характер и, наконец, зависимость их от состояний психической сферы, благодаря чему они колеблются в своей интенсивности вместе с переменами психического состояния.
Кроме психических анестезий и психических гиперестезии необходимо иметь в виду у душевнобольных еще разного рода
363
болезненные ощущения психического происхождения, которые, могут быть названы психическими алгиями или психоалгиями. Эти болезненные ощущения состоят в болевых и иных ощущениях в тех или других частях тела и во внутренних органах у душевнобольных, которые, находясь обыкновенно в тесном соотношении с психическим состоянием последних, отличаются большей частью блуждающим характером. Очень интересно, что у галлюцинирующих больных, как, например, при хроническом галлюцинаторном сумасшествии, эти психоалгии нередко вступают в теснейшее соотношение с слуховыми и иными галлюцинациями, возбуждаясь ими как бы по внушению и, с другой стороны, вызывая вслед за собой галлюцинации как явление последовательное. Особенно типичные примеры разного рода психоалгии представляют больные, страдающие хроническим галлюцинаторным сумасшествием алкогольного происхождения. Так, один из моих больных такого рода, помещавшийся в Казанской окружной лечебнице еще в бытность мою профессором в Казани и страдавший постоянными и обильными слуховыми галлюцинациями, подвергался самым разнообразным психоалгиче-ским ощущениям в виде уколов как бы шилом, жжений и пр. в тех или других частях тела.
Обычно психоалгические ощущения у этого больного сочетались с слуховыми галлюцинациями в виде слов: «клизмуем1 ревматизм!», так что каждый раз, когда больной испытывал то< или другое болезненное ощущение в определенной части тела, он тотчас же слышал от своих голосов заявление: «клизмуем ревматизм!», что, по его словам, обозначало, что эти вколы и жжения дело рук тех лиц, голоса которых больной слышал. Надо заметить, что у этого больного сфера чувствительности и движения не представляла никаких вообще объективных изменений и равным образом внутренние органы были здоровы.
У другой больной, страдающей хроническим галлюцинаторным помешательством с обильными слуховыми галлюцинациями и помещавшейся некоторое время в больнице св. Николая чудотворца, сложился бред, что врачи этой больницы во главе с главным доктором производят путем спиритизма разные непотребства и мучения или пытки над больными. Этот бред укрепился у больной под влиянием соответствующих слуховых галлюцинаций (слышание голосов врачей этой больницы), которые поразительным образом сочетаются у нее с разного рода психоалги-ческими ощущениями в наружных и внутренних частях тела. Больная сама подробно записывает свои ощущения и галлюцинаторные явления, вследствие чего лучшей характеристикой ее состояния могут служить хотя бы краткие выдержки из ее запп-
1 От слова клизмовать (т. е. делать клизму) — термин, вероятно,, введенный в употребление младшим медицинским персоналом Казанской окружной лечебницы.
364
сок, представляющих большое количество тетрадей, которые больная мне предоставила в надежде, что я ее избавлю от назойливых обидчиков.
От невралгических болей психоалгические ощущения отличаются тем. что они представляются чрезвычайно распространенными, тем, что часто поражают большим разнообразием в отношении своего характера и в то же время большой изменчивостью, тем, что находятся в прямой зависимости от психического состояния больных и, наконец, тем тесным соотношением с болезненными расстройствами сознания в виде галлюцинаций, с которыми мы познакомились выше.
О ГАЛЛЮЦИНАТОРНОМ ПСИХОЗЕ, РАЗВИВАЮЩЕМСЯ ПРИ ПОРАЖЕНИИ ОРГАНА СЛУХА
Уже давно известно, что поражения органов чувств приводят к развитию психических расстройств. Особенно благоприятную почву для развития таких расстройств представляют поражения органа слуха.
Имеется также целый ряд литературных указаний и на появление психических расстройств вследствие поражения органа зрения. Наконец, и поражения других органов чувств в известных случаях могут служить источником психических расстройств. Довольно нередки, например, по моим наблюдениям, случаи психических расстройств, развитие которых первоначально обусловливается поражением органа обоняния.
Что касается характера психического заболевания, обусловливаемого поражением органов чувств, то, судя по имеющимся в этом отношении литературным указаниям, здесь могут наблюдаться весьма разнообразные психические расстройства. Из них особенным разнообразием отличаются те психические расстройства, которые являются последствием поражения органа слуха.
Но из всех психических расстройств, развивающихся под влиянием поражения органов чувств, в клиническом отношении может быть выделена форма психоза, характеризующаяся главным образом более или менее стойкими обманами чувств преимущественно в области пораженного органа с вторичным развитием более или менее ограниченных идей бреда или даже при совершенном отсутствии последнего.
Таким образом, например, при хроническом поражении органа обоняния, психоз начинается обонятельными иллюзиями и галлюцинациями, к которым с течением времени присоединяются, по крайней мере эпизодически, и другие обманы чувств с характером большей частью иллюзий, что приводит обыкновенно к развитию идей бреда об издавании больным запаха (через задний проход, от мошонки, через рот и т. п.), который слышится будто бы также и другими, вследствие чего все от больного отворачиваются и т. п.. Подробнее об этой форме психоза мы скажем в другом месте; теперь же займемся лишь теми слу-
366
чаями, которые иллюстрируют развитие галлюцинаторного психоза при поражении органа слуха.
Надо заметить, что уже и в существующей литературе мы имеем несколько наблюдений с развитием обманов чувств в прямой зависимости от поражения органа слуха.
Так, например, Режи1 сообщает о больном, страдавшем правосторонними галлюцинациями с идеями преследования, зависевшими от гнойного отита правого уха. С другой стороны, Балль2 сообщает о больном, страдавшем гнойным катарром среднего уха с левой стороны, вследствие которого вместе с субъективными шумами в ухе развились также слуховые галлюцинации. Вилиус3 передает два случая, в которых развитие галлюцинаций должно быть поставлено в прямую связь с поражением органа слуха. В одном случае существовала otitis media hyperemia с обеих сторон, но справа выраженная резче, нежели слева. Вначале у больного с некоторым наследственным отягчением появился ушной шум, а затем и слуховые галлюцинации главным образом справа с последующим развитием бреда преследования. Местная терапия привела к благоприятному исходу заболевания. В другом случае у больного с местным отягчением развилось хроническое галлюцинаторное помешательство на почве существовавшей с детства otitis media purulenta duplex.
Имеется затем целый ряд других наблюдений, например, Мабиля4, Бюшерона5, Шмигелова0, Коццолино7, Ланнуа8, Балле9, Заухона10, Серпилли11, Белякова12 и др., в которых связь психических расстройств с поражением органа слуха представляется установленной с большей или меньшей точностью. Но при этом поражение органа слуха являлось этиологическим моментом
1 Regis, Des hallucinations unilaterales, L' Encephale, 1881.
2 Ball, Considerations sur un cas d' hallucinations de l'ouie conse-cutives a une inflammation chronique de 1' oreil moyenne, L' Encephale, 1881.
3 Wi 1 i us, Beitrag z. Lehre von der Reflexhalluzinationen, etc., Inaug.-Diss, Wiirzburg, 1883.
4 M a b i 1 1 e, Cas de guerison d' hallucinations unilaterales de 1' ouie de cause externe, Annales medico-psychol., 1883, t. X.
5 Bucheron, Folie melancholique etc., La France med., 1S87, t. II.
6 Schmigelow, Revue mens, de laringologie, 1887, № 8. 7Cozzolino,La psychiatria, 1887; реф. в «Архиве психиатрии»,
1889, т. XIII.
8 Lannois, Lesion de l'appareil auditif et troubles psychiques, Revue mens, de laringologie, 1887, t. VI.
9 Ballet, Hallucinations auditives etc., Annales medico-psychol., 1888, t. VI.
10 Sauchon, Uebcr einseitige Halluzinationen, Inaug. Diss., Berlin, 1880.
11 S e r p i 1 1 l , Riv. di freniairia. Реф. в «Neurol. Zbl.», 1890, N 2!.
12 С. Беляков, Вестник психиатрии, 1890, т. VIII, вып. 2. В статье этой приведена подробно и литература по вопросу о влиянии поражений органа слуха на развитие психозов вообще.
367
хорошо установленных и притом весьма разнообразных психических расстройств, выражающихся в одних случаях психическим угнетением, в других •— маниакальным возбуждением и даже явлениями, свойственными прогрессивному параличу помешанных.
Что касается вышеуказанной формы психоза, развивающегося под влиянием поражения органа слуха и характеризующегося обильными слуховыми обманами чувств с последующим большей частью эпизодическим развитием обманов чувств и в других органах, то больные этого рода часто сохраняют критическое отношение к своим галлюцинациям, в других же случаях высказывают на основании их бредовые идеи, объясняя происхождение своих галлюцинаций теми или другими посторонними влияниями.
Этот психоз, развивающийся большей частью у лиц, более или менее предрасположенных при поражении органа слуха, сопровождающемся теми или другими субъективными ощущениями, открывается иллюзиями и галлюцинациями в органе слуха обыкновенно неприятного для больного характера, причем обманы чувств в этом случае редко развиваются исподволь, чаще более или менее быстро, иногда даже сразу вместе с появлением или усилением субъективных шумов в ушах и держатся крайне упорно в течение продолжительного времени. Часто, наряду с неприятными галлюцинациями, проявляются безразличные по содержанию обманы чувств, а иногда и приятные; последние в отдельных случаях могут даже преобладать. Почти всегда галлюцинации усиливаются при обострении болезненного процесса в органе чувств, .сопровождающемся усилением и других субъективных ощущений, в другое время они более или менее значительно ослабевают. В отдельных случаях наблюдается даже совершенное прекращение галлюцинаций на известное время, что можно объяснить лишь улучшением болезненного процесса в органе слуха, но обычно, спустя то или другое время, слуховые галлюцинации снова появляются с большим или меньшим ожесточением. Нередко, кроме того, встречается одностороннее преобладание галлюцинаций.
Само собой разумеется, что больной вначале чувствует себя крайне смущенным появившимися обманами чувств и нередко приходит от них даже в ужас, проводя в волнении дни и ночи. Иногда он проявляет те или другие действия и поступки, руководимые обманами чувств, вступает в разговор с своими галлюцинаторными образами, бранится с ними, плюет на них, вообще обращается с ними, как с реальными существами, но затем, постепенно привыкая к последним, до некоторой степени оправляется от первоначального переполоха и овладевает собой в такой степени, что может заниматься своим делом, хотя мысли о причине новых явлений его не покидают. Начинается долгая продол-
368
жительная борьба между здравым рассудком, говорящим о невозможности реального существования явлений, соответствующих обманам чувств, и этими последними. В минуты появления обманов чувств больной, по крайней мере вначале, часто сам поддается мысли об их реальном существовании, дает ответы своим голосам и в свою очередь спрашивает их о том или другом, дабы выяснить для себя характер новых явлений. Как упомянуто, больной даже обнаруживает действия и поступки или делает заявление сообразно тем обманам чувств, которым он подвергается; но проходит некоторое время, обманы чувств ослабевают, и больной вновь начинает сомневаться в реальности своих обманов чувств. В этих колебаниях проходят недели и даже месяцы. Конечно, многое тут зависит от силы и характера самих галлюцинаций, степени интеллекта больного и, наконец, от большего или меньшего предрасположения его к душевным заболеваниям. Но обычно, если обманы чувств более или менее обильны и оживлены, к ним со временем присоединяются обманы чувств и в других органах, хотя бы эпизодическим образом; больные в конце концов поддаются своим обманам чувств и начинают так или иначе объяснять их происхождение, т. е. начинают давать ложное толкование своим галлюцинациям. Таким образом, дело доходит уже и до развития бредовых идей.
Но этот бред, большей частью основанный почти исключительно на обманах чувств, по крайней мере вначале, представляется несложным, однообразным и часто состоит в принятии реального существования галлюцинаторных образов и объяснения их происхождения и отношения к больному. При всем том этот бред, развивающийся большей частью не вдруг, а после долгой борьбы со стороны больного, не обнаруживает обыкновенно никакой склонности к прогрессированию и нередко даже в течение долгого времени представляется непрочно фиксированным, вследствие чего больные легко от него отказываются у даже поддаются убеждению в мнимости их галлюцинаторных образов, хотя большей частью только на время. В других случаях больные твердо убеждены, что они страдают галлюцинациями и признают их за болезненные явления, но в происхождении своих галлюцинаций обвиняют те или другие из посторонних влияний.
Упрочению и развитию бредовых идей способствует, конечно, всякое обострение обманов чувств, но особенную убедительность в глазах больных, как уже упомянуто, приобретает хотя бы временное появление обманов чувств в других органах, например, в органе зрения или обоняния. В этом случае большей частью дело идет об иллюзиях и галлюцинациях, развивающихся вторичным, т. е. психологическим, путем как прямое следствие обманов чувств со стороны первично пораженного органа или же на основании сложившихся предубеждений и тех или других бредовых идей. Во всяком случае эти обманы чувств в других органах играют лишь подчиненную роль в развитии пси-
24 В. М. Бехтерев
389
хоза. В конце концов у больных развивается бред о влиянии на них посторонних лиц при посредстве телефонов, электричества, телепатической передачи мыслей и слуховых ощущений и т. п.
Этот бред при условии, если поражение слухового органа не излечивается и обманы чувств не исчезают, продолжается обыкновенно в течение многих лет, иногда без всяких перемен, в других же случаях при осложнении болезненного процесса он со временем подвергается дальнейшему развитию. Исхода в слабоумие даже и по истечении многих лет я не видал ни в одном из своих наблюдений. Что касается выздоровления, то оно часто стоит в прямой зависимости от возможности устранить или улучшить основное поражение слухового органа, в противном случае болезнь тянется целыми годами, часто без всякой наклонности к дальнейшему развитию бредовых идей. Как упомянуто ранее, довольно нередко обманы чувств обостряются вместе с появляющимся в ушах шумом и шипением и даже слышатся в последнем; кроме того, в отдельных случаях наблюдаются даже односторонние обманы слуха, но в общем ход развития болезненных явлений мало изменяется от этих особенностей отдельных наблюдений. В отношении течения и ассоциаций идей, интеллекта и памяти у больных обыкновенно нельзя отметить ничего существенного. Настроение также не страдает первичным образом, вторичные же изменения его, вследствие обманов чувств, возможны и наблюдаются нередко по крайней мере в начале заболевания.
С физической стороны у больных находят обыкновенно указания на хроническое поражение органа слуха и более или менее заметное нарушение его функции, которое при тщательном исследовании вряд ли может ускользнуть от наблюдения; но в остальном почти никаких других явлений у больных открыть не удается. Что касается этиологии и патогенеза заболевания, то непосредственным поводом к его развитию служит всегда поражение органа слуха, чаще всего катарр среднего уха и евстахиевой трубы; при этом дело идет о поражении с характером воспалительного процесса, которое служит источником раздражения, приводящим к развитию субъективных слуховых явлений, за которыми обыкновенно уже и следуют обманы чувств.
Что касается неблагоприятной наследственности, то, пови-димому, она не может не играть известной роли в развитии психоза как предрасполагающий момент, но в иных случаях не отмечается никаких в этом отношении указаний и дело, вероятно, ограничивается личным предрасположением, выражающимся особой возбудимостью нервной системы, благодаря чему периферическое раздражение со стороны органа слуха поднимает возбудимость центров до появления в них болезненной реакции, выражающейся развитием иллюзий и галлюцинаций.
Представляя совершенно своеобразные особенности в клинических проявлениях и течении, рассматриваемый психоз,
370
по моему мнению, не может быть смешиваем ни с одним из общепризнанных психических заболеваний. От так называемой галлюцинаторной спутанности этот психоз отличается тем, что обманы чувств первоначально развиваются в одном из органов чувств и лишь исподволь присоединяются затем, и то большей частью эпизодические, обманы чувств и со стороны других органов; при этом никакой спутанности в сознании и никакого вообще расстройства в сочетании идей не обнаруживается. Самое течение психоза, а равно и развитие бредовых идей в том и другом случае совершенно иное. От хронического первичного сумасшествия рассматриваемый психоз отличается присутствием поражения органа чувств, обусловливающего заболевание, первичным развитием обманов чувств преимущественно с характером галлюцинаций и сосредоточением последних главным образом в одном из органов чувств, присутствием сознания своего болезненного состояния, часто даже критическим отношением к своим галлюцинациям, однообразием и относительной несложностью бреда, если он вообще имеется, бреда, стоящего вообще в прямой связи с упорством и стойкостью галлюцинаторных явлений, отсутствием первичных бредовых идей и отсутствием склонности к его прогрессивному развитию. Рассматриваемый психоз скорее всего может быть смешиваем по клиническим проявлениям с бредом преследования пьяниц (Нассе) или с чувственным бредом пьяниц (Крафт-Эбинг). В последнем, однако, галлюцинации хотя и обнаруживаются преимущественно в органе слуха, но большей частью проявляются одновременно и в других органах чувств и притом уже в самом начале болезни, приводя нередко к развитию сильнейшей реактивной тоски, при этом характер обманов чувств здесь принимает нередко крайне циническое содержание; вместе с тем идеи бреда здесь развиваются сравнительно быстро, большей частью с характером идей преследования, причем вместе с идеями преследования нередко появляются у больных и идеи величия или же идеи религиозного содержания. Кроме всего прочего, здесь имеются налицо характеристичные физические расстройства, свойственные хроническому алкоголизму, и соответствующий этиологический момент, обыкновенно открываемый без труда. Все эти данные позволяют выделить одно заболевание от другого. При этом должно иметь в виду, что спиртные излишества, способствуя заболеванию среднего уха, в известных случаях могут служить предрасполагающим моментом и к развитию рассмотренного нами психоза. Поэтому этот психоз развивается нередко у лиц, которые хотя и не могут считаться настоящими алкоголиками, но временно в большей или меньшей степени пользовались спиртными напитками чем и нажили себе хронический катарр среднего уха и носоглоточного пространства. При всем том несколько эксквизитных случаев заболевания я наблюдал у лиц, которые никогда не употребляли ни капли вина.
24»
371
Что касается предсказания, то, по крайней мере в начальном периоде болезни, оно стоит в прямой связи с возможностью излечить или улучшить основное поражение, а потому все внимание врача должно быть направлено на исправление существующего поражения органа слуха. К сожалению, это далеко не всегда возможно, так как обычно мы имеем здесь дело со старыми болезненными процессами, мало поддающимися излечению.
В заключение приведу для иллюстрации нижеследующие наблюдения. Других, относящихся сюда же наблюдений я не привожу, так как хотя они и не менее характеристичны, но не все из них отличаются должной полнотой, так как такого рода случаи по самому характеру заболевания чаще подвергаются амбулаторному наблюдению, нежели клиническому или больничному.
Наблюдение 1. Офицер Ш., около 40 лет. В семье нервных и душевных болезней не было. Сам больной с детства был нервным, но особыми тяжелыми недугами не болел. Спиртными напитками никогда не злоупотреблял. Пьет только по рюмке за обедом и за ужином.
Около месяца тому назад после большого переутомления и бессонных ночей при шуме в ушах у него появились слуховые галлюцинации, которые сам больной описывает следующим образом:
«Я женат уже около 15 лет. Дети жены от первого брака жили почти все время (кроме настоящего 1902 г.) у наших родных, так как мне приходилось служить в городах, не имеющих подходящих учебных заведений.
В 1902 г., по переводе меня на службу в Минск, мы взяли детей к себе. Все время нашей совместной с детьми жены жизни все шло отлично без неприятностей.
Вследствие усиленных летних и осенних занятий нервы мои сильно расстроились и я иногда не мог заснуть, несмотря на физическое утомление, часов до 2—3 ночи. Числа 25—26 сентября с. г., в одну из бессонных ночей мне вначале послышался стук часов (будильника), но, прислушиваясь дальше, я к удивлению начал разбирать слова и фразы. Принял я эти слова за стук часов только вначале, когда темп произношения их подходил к стуку часов, но затем фразы стали произноситься более протяжно (с целью, вероятно, чтобы я мог уяснить себе смысл слов и фраз) и я услышал, что слова походили на звуки, производимые шуршанием о спичечную или картонную коробку. Я удивился этому и перестал обращать внимание и, кажется, заснул.
Днем ничего я еще не слышал. Дня через два перед сном я вновь услышал слова и фразы и усмехнулся их содержанию. Тогда я услышал вопрос: «Чему я смеюсь?». Я удивился и подумал, кто это мне говорит и почему он знает, что я засмеялся. На это я тоже получил ответ, что говорит мне это такая-то (падчерица), а что я смеялся — это она узнала, но как ■— не может и сама объяснить этого.
Ввиду того, что комната падчерицы отделяется от нашей спальни залой, столовой и комнатой пасынка, я невольно подумал, каким способом могу я слышать фразы на такое расстояние и с кем она переговаривается, но и на это я получил ответ, что разговоры ведутся ею с ее братом посредством шороха о коробку, а что мои мысли (так как я не знаю этого способа разговора) она узнает тоже, но не знает почему. Тогда, подумав о том, что падчерица моя была особа малокровная и нервная (от чего и лечилась раньше), я пришел к тому убеждению, что и я расстроил себе нервы так, 'что подошел к уровню ее нервной системы и наши нервные системы нашли точку соединения в мысленном разговоре на расстоянии.
Заинтересовавшись этим и желая проверить, я стал мысленно задавать ей вопросы и, думая, что, может быть, задавая вопросы, я сам же составляю и воспроизвожу себе ответ на него, я стал задавать такие во-
372
просы, ответов иа которые я не мог бы составить себе сам, не зная ни обстановки, ни тех обстоятельств, прикоих мог создаться ответ, и на эти вопросы я получал верные ответы (так как я проверил их на деле, осторожно расспрашивая других лиц). Напротив того, на те вопросы, иа которые я мог. ответить, голос отзывался незнанием; слова мои (выражения технические и т. п.), которые я произносил скоро, голос перепутывал, а если и повторял, то медленно и раздельно. Начавшись с этого вечера, разговоры эти велись затем и днем, но изредка и по уходе моего или падчерицы из до-' ма — прекращались. Вне дома я ничего не слыхал, да и дома слышал вопросы только тогда, когда читал какую-либо книгу. В этом случае голос спрашивал значение некоторых слов, выражал одобрение или неудовольствие читаемому, иногда то, что мне было интересно, как специально военное, ему не нравилось, и наоборот, — одним словом голос всем стал интересоваться. Сначала это занимало меня (я даже разъяснял голосу требуемое им), но ввиду того, что это отвлекало меня от дела, мне стало надоедать и я просил оставить меня в покое. Голос вначале извинялся и обещал прекратить разговоры, но через некоторое время с легкомыслием женщины вмешивался вновь. Так было дня четыре, как днем, когда я был дома, так и вечером. Но затем (я думаю потому, что все что я делаю, делаю осмысленно, т. е. беря, например, какую-либо вещь, я в то же время и соображаю, хотя и бессознательно, что я беру), я стал уже слышать голос, говорящий мне, что я делаю, или о чем думаю в настоящий момент,, и меня начало уже тяготить сознание того, что я становлюсь уже не полным хозяином своих мыслей и своих действий и что все это становится достоя-, нием и другого лица. Это меня и убивало, и раздражало в то же время. Я вначале просил прекратить это ( я уже помирился с тем, что все это факт),, я затем, раздражаясь, ругал. В конце концов и на меня посыпались ругательства. Вопросы чередовались, когда я не обращал на них должного внимания, с ругательствами по моему адресу. Раздражаясь день ото дня, я, наконец, не выдержал и, думая, что фразы я слышу шорохом по предмету (шорох был очень силен и назойлив для моих ушей), прошу жену послушать, что мне говорят. Жена вначале прислушивалась и сказала мне, что не слышит ни голоса, ни шороха; это меня еще больше удивило и в то же время я пал духом. Звуки после этого продолжались еще чаще и слышнее, ругательства усилились. Я просил оставить меня в покое, что я не желаю им зла и что нечестно пользоваться этим для ругательств и тому подобных мерзостей, но голос мне заявил, что оба они (т. е. пасынок и падчерица) ненавидят меня и доймут.
Я решил обратиться к докторам и они нашли, что у меня развилось малокровие, болотная лихорадка и неврастения со слуховыми галлюцинациями. Успокоенный я вернулся домой, рассказал о мнении доктора и решил лечиться и не обращать внимания на донимающий меня голос. Сидя в этот вечер в зале за книгой, я вновь, услышал голос, но, следуя совету врача, не стал обращать внимания. Голос забеспокоился о том, что, кажется, доктор убедил меня в галлюцинации слуха. Повелись с кем-то вдвоем переговоры об этом (причем слово галлюцинация произносилось растяжимо). Затем голос стал убеждать меня, что доктор не прав, и в доказательство того, что все это правда, а не галлюцинация, голос заявил, что пасынок сейчас пройдет через залу в переднюю, что и проделалось тотчас же, а когда я не поверил и этому и счел это за случайное совпадение, то такое фактическое убеждение повторилось в этот же вечер и в другой раз. Я уж и не знал, что мне и думать обо все этом. Факты брали свое. Дня два я лечился, но голос и днем, и вечером донимал меня. Я просил и пасынка, и падчерицу сказать правду; я прощаю им, если они и ругали меня, не беда! Но мне нужно знать, правда это или же я сильно болен; просил сказать правду хоть не ради меня самого, а ради матери их, которую все происходящее со мной убивает. Но они говорили мне, что я, вероятно, сильно болей и что они не только меня не ругали, но что и не за что меня им ругать. Их лица были спокойны и только выражали любопытство. Я сбился совсем с толку и просил простить меня за беспокойство.
373
Но лежа в этот вечер, я вновь слышу ругательства; я не обращаю внимания, а они мне говорят, что они никогда не сознаются, так как боятся матери, меня же они дурачат и доймут. Решив проверить еще, я прошу жену поместить детей на время у знакомых. В тот вечер, когда они были у знакомых, я вначале не слышал ничего, но когда я решил исследовать, не происходит ли воображаемый мною голос от каких-либо других причин: от стука будильника (в кухне), шума вентилятора в столовой или другого какого-либо шума, происходящего вне дома и концентрирующегося в комнатах, для чего я решил пойти в комнату пасынка и прилечь на его кровать, то как только я подумал это сделать, как услышал отдаленный едва разборчивый голос, запрещавший мне делать это. Пройдя в столовую, я ясно услышал шум вентилятора и стук часов, но слышал в то же время и голос (очень слабо). Голос слышал я именно с той стороны, где находилась квартира знакомых. В этот же вечер я получил толчок в левое плечо, причем голос сказал мне, что мне нанесен удар в плечо, затем удар в голову. Удары были слабые, но удары в голову имели последствием слабый же звон в ушах. Тогда я и сам стал наносить мысленно удары и услышал просьбу не ударять так сильно, причем указывалось и место удара. Дети жены были возвращены в дом, так как не имело смысла держать их особо.
Через несколько дней я узнал еще особую странность: мне казалось иногда, что я ем (мои челюсти проделывали процесс еды и я чувствовал даже нажим концами зубов на что-то твердое) и даже глотаю, хотя, глядя в зеркало, я видел свое лицо спокойным. Я недоумевал, но голос сообщал мне, что субъект, говорящий со мною, кушает яблоко. Это уже было днем.
Все эти вещи повторялись затем и днем; и голос, и удары я слышал сильнее, когда падчерица была ближе ко мне, и слабее, когда дальше, — расстояние ослабляло их.
Здесь, в Петербурге, и в дороге я не слышал и ие слышу ни голоса, ни ударов. Во всем остальном я чувствую себя совершенно здоровым и память у меня довольно сильна».
В дополнение к сделанному описанию замечу, что больной, первоначально думавший, что дело идет о передаче мыслей на расстоянии, в настоящее время вполне убедился, что это галлюцинации, вследствие чего и пришел ко мне за советом.
С физической стороны ничего патологического; рефлексы в порядке, шума в ушах в настоящее время нет; исследование же, произведенное проф. Верховским, показало, что у больного имеется хронический насморк, хронический же катарр носоглоточного пространства и боковой двусторонний фарингит.
Наблюдение 2. Больная И. поступила в клинику 7 октября 1902 г. Ей 42 года, девица, рясоформная монахиня общежительного монастыря в Калужской губ. Наблюдение о ней составлено д-ром Певницким. Больная худощава. Телосложение правильное. Внутренние органы без заметных уклонений от нормы. В ушах гиперемия молоточка и cavi epitympanici. Незначительное отклонение языка вправо. Болевая чувствительность несколько острее слева. Рефлексы со слизистых оболочек нормальны, сухожильные—повышены равномерно; сужения поля зрения нет. Кожные рефлексы повышены. На пальцах обеих ног вросшие ногти>
Больная из духовного звания. Через год после ее рождения отец ее умер от скоротечной чахотки, а мать умерла позже, на 41 году жизни. Брат отца после неудачного романа был душевнобольной, но исхода его болезни больная не знает. Брат матери — протодиакон, много пил.
Больная развивалась правильно, но никогда не отличалась особым здоровьем. Училась хорошо. Окончила епархиальное училище и была учительницей 12 лет; затем она, наезжая к своей родственнице в монастырь.
374
привыкла к монашескому обиходу и, послушавшись советов одного монаха, осталась в монастыре, где пробыла 12 лет.
Регулы у больной начались в 13 лет, установились в 16 лет. В этом жг году У нее был роман, который не окончился браком, так как больная боялась брачной жизни. Она, повидимому, давно и постоянно мастурбирует, по крайней мере в ее записках имеются жалобы на то, что приятные половые ощущения теперь получаются труднее, так как на половых губах образовались ссадины. На расспросы о своей половой жизни она отвечает уклончиво. Спиртных напитков больная вообще никогда не употребляла. В своей жизни больная много болела; у нее был тиф, воспаление легких, дифтерит. Она страдала геморроем и сильными маточными кровотечениями, которые были на 37 году жизни и сопровождались изменениями психики: больная была очень раздражительна и все ей были противны.
Ее жизнь в монастыре была тяжелая: ее усердно эксплоатировали. Сначала она была учительницей, потом занималась по счетоводству и выбивала по золоту. Работы было много, отношения монахинь скверные, кормили скудно и однообразно.
Десять лет тому назад больная простудилась, у нее развилось воспаление ушей, произошло по ее словам омертвение барабанных перепонок, появились боли в ушах, и она потеряла слух. Заболевание продолжалось месяц, барабанные перепонки зарубцевались, осталась лишь вздутость их, причем слух возвратился, но не вполне. Через 11/2 года наступил возврат заболевания, еще через год — второй, и слух стал значительно ухудшаться, слева больная даже совершенно оглохла. Появились постоянные шумы в ушах.
Вслед за развитием ушных шумов при выбивании по золоту больная вдруг стала слышать в стуке свое имя и отчество. Затем она вскоре услышала целый разговор между ней и девочкой, которую она когда-то наказывала в школе. Затем разговоры стали слышаться все чаще и чаще, например, она слышала площадную брань, когда стояла в церкви. Когда она сообщала об этом игуменьи, та увидела в этом бесовское навождение и приказала ей больше молиться. Но чем больная усерднее молилась, тем ругань становилась циничнее. Больная перестала ходить в церковь и отправилась к архиерею с просьбой выпустить ее из монастыря. Архиерей приказал ей дожидаться ответа в своем монастыре. Больная вернулась обратно и вскоре была игуменьей в сопровождении двух монахинь отправлена будто к архиерею, а на самом деле в психиатрическую лечебницу. Это обманное помещение сильно взволновало больную; ее через месяц выпустили. С месяц голосов совершенно не было вместе с прояснением слуха, а затем они вернулись с большей силой при новом ухудшении слуха. Временами голоса обостряются, их особенно много было при посещении больной о. Иоанна Кронштадского, что, вероятно, также следует объяснить ухудшением слуха, вследствие продолжительной дороги.
В настоящее время больная ведет себя спокойно, много и охотно работает, но часто нервничает, так как ее донимают галлюцинации. Она постоянно слышит разговоры знакомых и окружающих, шумы, стуки, хрипение. Все это слышится ей там, куда направлено ее внимание. Стоит ей почесаться, например, и прислушаться, она и с места чесания слышит голоса. При засыпании в месте прикосновения лица к подушке ей слышится разговор в последней. Слышит она чаще всего не разговоры, а отдельные фразы или слова, например, «Катенька (имя больной) перевернется» или «к доктору привернулась» и т. п. Эти фразы или обрывки фраз повторяются и этим раздражают больную. Но иногда больная слышит и целые разговоры, которые она не передает устно, а записывает. В них ясно отмечается эротический оттенок; голоса или предлагают ей войти в половые сношения, или упрекают ее в этом. Кроме того, она различает чужие голоса, голоса далеких знакомых и окружающих. Голоса эти слышатся и в ней, и на ней, например, в груди и на груди, в носу, придыхании, во рту. Далее, ей чудятся изредка разные противные запахи, между прочим, запах гноя костоеды; иногда же чувствуется отвратительный вкус гнилья и щипанье языьа
375
во рту, в другое же время — прикосновение мертвеца или поцелуй ее руки кчким-то знакомым. Язык ее иногда сам двигается или как будто бы кто-то его двигает (психомоторные галлюцинации). Все эти явления больная обычно не признает за реальные явления, но она убеждена, что в этом проявляется злая воля игуменьи Ефросиний и купца П. На игуменью она подала даже жалобу обер-прокурору Св. Синода, где описала все свои галлюцинации. По мнению больной, П. снял в граммофоне ее голос и голоса и речи других, знакомых ей, и какими-то невидимыми нитями, вероятно электричеством, передает ей эти разговоры. Эта масса галлюцинаций временами настолько подавляет больную, что она раздражается и заявляет, что если не поправится, то она повесится. Больная легко гипнотизируется, получается 3-я стадия по Льебо с амнезией. Легко вызываются каталептические явления, автоматизм и послегипнотичсские внушения, но голоса путем внушения устраняются лишь на несколько часов. Исследование ушей, произведенное специалистом, показало ясные следы хронического катарра обоих средних ушей.
Наблюдение 3. Больная X., около 25 лет, происходит из семьи, в которой брат был душевнобольным, отец злоупотреблял спиртными напитками. Сама больная всегда отличалась удовлетворительным здоровьем. С год тому назад она имела роман, окончившийся отказом с ее стороны выйти замуж, около полугода назад у ней развился обоюдосторонний otitis medialis, причем справа, вследствие гнойного процесса, потребовалась трепанация сосцевидного отростка. Слух восстановился, но начали случаться с тех пор сильные головные боли и остались шумы и свисты, особенно сильные справа. Недели 2 спустя после трепанации у больной появились впервые галлюцинации слуха. Она начала слышать несколько голосов, раздающихся извне, в числе которых немаловажную роль играл и голос любимого ею господина. Эти голоса продолжаются и по-'сие время, причем они то неприятны для больной, то приятны. Наряду с голосами, у больной имеется и род неполных галлюцинаций (псевдогаллюцинаций) в виде «громкого мышления». Голоса больная слышит правым ухом, в котором делалась трепанация сосцевидного отростка и в котором по настоящее время слышится сильный свист и шум, тогда как в левом ухе субъективные шумы хотя и имеются, но много слабее. Иногда голоса слышатся как бы в шуме и свисте. Галлюцинаций и иллюзий в других органах не имеется. Больная признает, что она страдает галлюцинациями и благодаря этому очень рассеянна; но полного критического к ним отношения, повиди-ыому, не имеет, вследствие чего по временам она склонна поддаваться слуховым галлюцинациям и под влиянием голосов говорит со своими родными о нарядах и о богатстве, что совершенно не соответствует их бедной обстановке. Исследование обнаружило мутность и втянутость барабанных перепонок и вероятное их приращение к стенке среднего уха.
Наблюдение 4. Больной М. обратился за советом ко мне еще в 1886 г., он офицер, по происхождению дворянин, Кавказского племени, магометанского исповедания, 38 лет, представляет собой субъекта несколько выше среднего роста, хорошего телосложения. Заболеваний внутренних органов нет. Страдает катарром средних ушей, особенно выраженным с правой стороны. Из аномалий развития можно указать на существование особой пигментации в виде пятен на правом райке и на широкое расстояние между средними верхними резцами.
Отец больного во время зачатия последнего был под сильным нравственным давлением из-за неприятностей по службе. Вообще об отце можно сказать, что он был умный и дельный человек; временами покучивал. Мать была очень умная женщина; часто страдала головокружениями и головными болями. Дядя по отцу был подвержен запоям. Сестра больного унаследовала от матери головокружения и шум в ушах.
Сам больной в течение всей своей жизни перенес несколько болезней: в детстве была сильная простуда, от которой лечился около 6 месяцев; в детстве же раз опрокинул на себя горячий самовар, обварился и сильно испугался, около 7—8 лет от роду упал в кустарниках и потерял сознание,
376
после чего было лихорадочное состояние; около 15 лет от роду перенес тиф. Как запомнит себя страдает приступами головных болей, продолжающимися 1—2 дня через неопределенные периоды времени. Во время этих приступов, кроме болей в голове, всегда были какие-то неопределенные ощущения особого характера. Облегчение от приступов являлось всегда вслед за выделением из носу. Всегда больной отличался раздражительностью из-за пустяков: сердился и ссорился часто без уважительных причин, хотя в общем он очень интеллигентный человек. В компании с товарищами иногда позволял себе немного выпить и при этом замечал за собой, что от водки быстро являлось возбуждение, сменявшееся быстро же озлоблением и затем угнетением, причем больной уходил из компании. Вследствие этого больной избегал алкоголя и злоупотребления последним у него вообще никогда не было. По его рассказу, болезнь началась следующим образом:
В январе 1885 г. больной должен был ехать из Севастополя в Петербург, чтобы поступить в школу. Накануне отъезда больной кутнул с товарищами. Не будучи из пьющих, он во время езды по железной дороге ничего не пил и только в Курске выпил одну рюмку. На одной из станций между Курском и Москвой около 10 часов вечера больной вышел на вокзал и выпил стакан чаю; затем вошел в вагон 3-гр класса, в котором ехал, и сел, полулежа и облокотившись. Спать не хотелось. Около него сидел очень солидный человек с женой; затем сидел хорошо одетый еврей и неподалеку денщик больного. Вдруг больной слышит, что солидный человек говорит еврею: «Я видел много военных, но такого фата, как этот, не видывал; видишь, как грудь выпятил; должно быть набитый дурак!». За разъяснением этого больной обратился к еврею, но тот не дал обстоятельного ответа. Затем больной заметил, что некоторые пассажиры стали переменять свои места, толкались друг с другом и перешептывались между собой. Позади больного ехал один пассажир с женщиной; на руке этого пассажира больной заметил кольцо, женщина же вдруг произнесла: «этакий чертенок; так бы я его искусала!». К кому относились эти слова — больной не знает. После этого женщина легла спать на скамейке позади больного. Вдруг он чувствует, что она, перекинув руку через перегородку, схватила его за голову; он ясно ощущает прикосновение ее руки к своей голове, по ощупать рукой он не догадался, так как напал на него какой-то необъяснимый страх и выступил холодный пот. Затем эта женщина говорит: «Да-лай 100 рублей!». Видя, что я ничего не отвечаю, она просит 50 рублей, затем 35 рублей. В это время в другом конце вагона слышится сильный шум голосов двух или трех человек; голоса выкрикивали «страшную па-хабщину». Под влиянием этого крика больной, уставши, уснул.
Поутру, проснувшись, выпил чаю и умылся; голосов не было. После того, как сел в вагоне, снова явился голос, рассказывавший всю биографию больного с его рождения. Это был сильный басистый голос, похожий на голос одного его знакомого, часового мастера в Петербурге и поэтому ему казалось, что это говорит тот самый часовой мастер. Далее голос говорил, что, хотя больной едет в Петербург для поступления в школу, но он не поступит, потому, что один командир — враг больному, живет с его женой и воспрепятствует поступлению. Больной ужасно растерялся и не знал, что делать. В это время он посмотрел на сидевшего вблизи «солидного человека», который вчера рассуждал о больном с евреем, и увидел, что у него являются глотательные движения и при этом «движения кадыка». В один миг в голове явилась мысль, что это был «чревовещатель». При этом больной услышал еще голос будто бы этого самого «чревовещателя»: «Хорошо не узнавать своих знакомых!». И тотчас ему представилось, что это, действительно, знакомый человек. Затем этот человек ушел в другую половину вагона.
На одной из станций в вагон село несколько новых пассажиров. Один из пассажиров сказал денщику больного: «Эй, ты, крупоизвергательН Пассажиры больному показались пьяными, так что на следующей станции он обратился к начальнику, чтобы тот запретил бесчинство.
377
Потом больной слышит, что «солидный» знакомый «чревовещатель» говорит: «Экий невежа!» Снова на станции сели новые пассажиры и голос начал говорить, что жена его здесь, племянник здесь и здесь же сестра его жены и некоторые другие его знакомые. Одна старуха показалась ему знакомой, но замаскированной; голос сейчас же и замечает, что это такая-то знакомая. Затем голос говорит, что жена больного умерла, его это встревожило, он расплакался, лег и уснул. Проснувшись, он услышал голос одной пассажирки, похожий на голос его родственницы. Тотчас же ему представилось, что это, действительно, та родственница, на голос которой похож голос пассажирки, и он подошел к ней спросить об этом, чтобы окончательно увериться. Вслед за этим больной вышел на площадку вагона и хотел броситься, но раздумал и ушел в другой вагон. Однако и там слышались те же голоса из первого вагона, доносились издали. Теперь голоса стали говорить даже о таких эпизодах жизни больного, о которых он и сам едва ли когда-либо думал.
По приезде в Москву больной вышел на вокзал и спросил себе стакан чаю. За чаем слышит голос: «Здесь татары содержат буфет, поэтому не надо платить денег». Лица содержателей, действительно, похожи были на татарские. Чтобы удостовериться, нужно ли, действительно, платить за чай, больной спросил о цене и получил в ответ: «15 копеек за стакан чаю». В другой группе пассажиров слышатся голоса: «Одурачим этого дурака!». На Николаевском вокзале в Москве больному все представляются его знакомыми и голос подтверждает, что это, действительно, его знакомые. Одна по костюму была совершенно схожа с одной родственницей больного, но когда он ближе стал разглядывать ее лицо, то оказалось, что это совсем чужая. Затем голос говорит: «ты не поезжай с этим поездом; для тебя будет особый поезд». В ожидании поезда больной подошел к буфету спросить стакан чаю и тут видит ехавшего с ним «солидного» господина, который будто бы готовитпоезд для больного. Однако этот господин должно быть заметил необычный вид больного, встал и ушел. Затем — поезд готов. Вдруг в это время слышится шум, музыка, и голос стал говорить, что это «его встречают». После того как больной сел в вагон, голоса опять начали рассказывать эпизоды из его жизни. Затем вдруг в одном углу вагона он видит 2-летнего мальчика в ермолке, которого одна женщина то поднимает вверх, то опускает; левый угол рта у этого ребенка поражен раком и лицо его постоянно «менялось». Такого же ребенка, только девочку, больной видел еще в детстве, но никогда до этого случая не приходилось вспоминать об этом.
В Твери садится против больного одна довольно приличная женщина и при этом замечает: «С вами мне будет веселее». Разговорились, больной предложил компаньонке подушку, на которую она и легла, продолжая беседу с больным. Когда умолкли, вдруг больной слышит сзади крики: «Яу-ну, расправляйся!». Пассажирка этим как будто сильно была обескуражена. Больному затем начал говорить голос его жены, которой тогда тут не было: «Перейди в спальный вагон!». Но больной хотя и был утомлен, однако не перешел на этот раз, перешел он лишь через несколько станций и спал хорошо. Утром голоса с самого утра начали преследовать больного и укоряли его: «А! Когда мы советовали перейти в спальный вагон, тогда ты не перешел, а теперь перешел!». Голоса усиливались и их становилось все больше и больше. Раздавался какой-то крик. Голоса стали слышаться уже в шуме колес, в скрипе их и по стенам вагона. Представлялось, что несколько человек ложилось лицами к стене вагона и пускали разные ругательства по стене; эти ругательские голоса, расходясь, усиливались. Чтобы удостовериться в истинности всего совершающегося, больной предложил прислушаться у стены одному своему товарищу, на что тот ответил, что шум, действительно, слышеи, но голосов никаких нет и, конечно, был удивлен вопросом.
После остановки на одной из станций больному захотелось есть; вдруг он слышит голос: «Не пей водки!». Не хотел пить, но тут на зло спросил 2 больших рюмки н выпил; после этого вскоре, утомившись, заснул.
373
.Через несколько станций голоса опять говорят: «Выпей водки!» Тогда на зло же им не выпил.
Рядом, в дамском отделении спального вагона, ехала одна дама. Она однажды стала у двери и смотрела на больного. Тогда голоса начали страшно ругать больного и ругали так, что, как ему казалось, могла все слышать эта дама. Больному было очень стыдно; но вскоре голоса стали хвалить больного, а барыне не советовали засматриваться на него, потому что она может влюбиться и тогда уже ей не вернуться в Туркестан, откуда она ехала. После этого вскоре барыня скрылась и более не показывалась, но голоса стали ругать и барыню.
По приезде в Петербург больной вышел из вагона, взял извозчика, приехал в гостиницу, причем голосов более уже не было; вместе с тем прекратились и ушные шумы.
Но однажды в июне больной ехал в вагоне конки и слышит: «Фуй, какая дрянь!». На это больной, однако, не обратил достаточно внимания. Затем в первых числах июля 1885 г. больной был на мусульманском празднике в Петербурге. В первый день праздника он был очень утомлен, но к вечеру при всем том вместе с женой отправился для прогулки в сад, откуда они вернулись на ночлег домой только около полуночи. Жена больного любила особенно спорить относительно семейных добродетелей жен; при этом она отличалась нетерпимостью к увлечениям жен и находила даже обидным для женщин снисхождение к таким женам со стороны мужей. Во время прогулки в описываемый вечер у больного с женой был этот же обычный спор. Больному на этот раз очень не нравилось то, что в суждениях жены проглядывала какая-то «особенная самостоятельность», которая ему казалось подозрительной. По возвращении домой легли спать. Затем жена закашляла и кашлянула 8 раз. Больному этот кашель показался очень подозрительным. Он не мог заснуть, жена тоже не спала Через несколько времени больной перешел на диван, где и заснул. В 5 часов утра жена опять закашляла и опять кашлянула 8 раз. В 8 часов утра больной слышит шаги около окна на дворе, затем обратно шаги и с «условным» кашлем. В это время жена встает, отдергивает занавеску и смотрит туда. С этого времени у него опять начались галлюцинации. Подозрительность по отношению к жене явилась страшная и он всячески старался выведать скрывавшуюся от него тайну. Раз после этого он слышит голос в печную трубу снизу вверх: произносится имя и отчетство его жены. Больной слышал этот голос, притаившись. Это был голос, не похожий на другие голоса, и больной считает его реальным. Подозрения по поводу неверности жены усиливались. Среди таких размышлений раз ночью, около 11 часов, при свече больной слышит голос: «Хочешь видеть свои рога?». Отвернувшись, он увидел на стене большие рога, как у оленя, которые скоро, как стал пристально всматриваться, исчезли. Однажды больной забыл принять назначенные ему для лечения пилюли, вспомнив о них позже времени приема, за чаем. Тотчас же голос и говорит: «Назначено натощак, а ты за чаем!». В другой раз больной должен был прочесть публичную лекцию и боялся за ее удачу, но говорил, шутя, что, если лекция выйдет неудачной, то он сошлется на свои голоса. Сейчас же голос и говорит, что «они об этом ему не говорили». В другой раз было так, что больной должен был прекратить из-за голосов лекцию, потому что чем дальше продолжал ее, тем голоса все более и более усиливались.
Чем больше бывает взволнован больной, тем голоса сильнее и сильнее. При размышлениях о неверности жены голоса на этот счет еще более надоедают больному. Голоса затем являются часто вроде передаточного пункта между больным и женой. Что нужно передать жене, то передают голоса и больной слышит, а что нужно передать ему, то он все понимает. Бывает, кроме того, и такое состояние: если больной будет глотать слюну, то ему представляется, что он не глотает, а жена, и голоса говорят, что жена глотает слюну, а не он. О чем бы больной ни подумал, о том появляется целый разговор. Иногда же голоса идут в разрез с мыслями больного, но они могут быть направлены и по желанию больного. Иногда раз-
379
говоры других понимаются не так, как они говорят, а как направляют мысли больного голоса. Последние слышны иногда в капанье воды, в плеске ее, в скрипе колес и т. п., но в это время уже голоса со стороны не слышны. Вместе с действительными звуками локализация голосов нередко изменяется. Они начинают выходить из места, соответствующего источнику звука. При прислушивании голоса сильнее, как и при волнении; при покойном состоянии голоса затихают. При занятиях и увлечениях голоса часто совсем не слышны и во всяком случае они меньше. На улице и дома голоса одинаково беспокоят. Чем дальше они раздаются, тем они громче преследуют больного. В пустом пространстве всегда слышны издали. При закрывании уха голоса так же слышны, как и с открытыми ушами. Они всегда слышнее справа (со стороны более пораженного уха), чем слева. Голоса во 2-й раз их развития происходят на татарском языке, а первый раз они слышались только по-русски. Больной совершенно критически относится к своим голосам, но первое время был уверен, что голоса слышатся не им одним, но и другими, от чего затем отказался после убеждения со стороны других.
Некоторые из голосов, по заявлению больного, несомненно, представляют повторение его мыслей, как бы «громкие думы». Другие голоса находятся в противоречии сего мыслями и отличаются необыкновенной объективностью. Иногда голоса говорят: «Не ешь отравленное!». Галлюцинации со стороны других органов чувств вообще крайне редки. Раз на лекции больного кто-то схватил за плечо. Раз ночью, когда больной лег спать, он ясно слышал запах мускуса. В другой раз ощущал запах дыма,как будто что-то начинало тлеть, и в это время слышал крик «пожар!». Затем представилась лунная ночь, туман, ветер и напевание голосов: «Кумушка, голубушка». Вот и все, что может припомнить больной, кроме упомянутых выше «оленьих рогов».
Иногда больной не может дать себе отчета — свои ли мысли у него происходят или он слышит чужие голоса. Во всяком случае больной относится вполне критически ко всем описанным обманам чувств; у него не наблюдается никакого вообще бреда и душевная деятельность в других областях не представляет заметных уклонений. Больной очень интеллигентный человек, занимающийся хозяйством со счетной частью при своем полку.
Исследование ушей, произведенное специалистом, обнаружило присутствие otitis media simplex bi lateralis, выражающееся мутностью, одинаково выраженной на обеих барабанных перепонках; имеется также и втянутость барабанных перепонок, выраженная на левой стороне сильнее, чем на правой. Подвижность барабанных перепонок представляется одинаковой. Обе евстахиевы трубы проходимы. Имеется, кроме того, старый ка-тарр носоглоточной полости. На правое ухо слух обнаруживает умеренную степень гиперестезии. Продувание не обнаруживает влияния ни на субъективные ощущения, ни на галлюцинации.
О НАРУШЕНИИ ЧУВСТВА ВРЕМЕНИ У ДУШЕВНОБОЛЬНЫХ
Чувство времени за последние два десятка лет сделалось, как известно, предметом многочисленных экспериментально-психологических исследований и в этом отношении оно может считаться наиболее изученным из всех так называемых центральных чувств, расстройства же этого чувства, наблюдаемые у душевнобольных, до сих пор, повидимому, мало обращали на себя внимания; между тем в отдельных случаях они, несомненно, играют немаловажную роль в общем симптомокомгшексе душевных заболеваний. Ввиду этого весьма желательно, чтобы на эти своеобразные расстройства у душевнобольных было обращено больше внимания, чем это уделялось до сих пор.
Чаще всего нарушения чувства времени у душевнобольных выражаются тем, что оно является в такой степени ослабленным, что больные при отсутствии какого-либо помрачения сознания совершенно лишены возможности ориентироваться во времени и не могут более или менее точно определять, сколько прошло времени от того или другого известного им события, несмотря на то, что память их вообще и в частности локализация бывших до болезни событий не представляется существенно нарушенной. Это более обыкновенные случаи нарушений определения времени, наблюдаемые у душевнобольных довольно нередко. Очень возможно, что они могут быть объясняемы наступившим с болезнью ослаблением способности узнавания прошлых представлений, с которой связана и наша способность локализации представлений во времени.
Другие, уже более редкие расстройства чувства времени выражаются тем, что больные значительный, иногда даже огромный промежуток времени уменьшают до минуты, в других же случаях минута им кажется вечностью. Чтобы иллюстрировать это состояние примерами, я остановлюсь на нескольких из своих больных.
Больной И. С, купец, алкоголик, ежедневно употреблявший водку по нескольку рюмок за столом и, конечно, не признающий себя алкоголиком. О ближайших родных подробностей известно мало.
В июле месяце 1901 г. после продолжительного купания заболел психически, впав в угнетенное состояние. Вскоре это угнетение достигло
381
своего апогея. Больной проявлял постоянное беспокойство, охал, стонал, бродил бесцельно по комнатам, ни с кем не говорил, обнаруживал недоверчивое отношение ко всем и, очевидно, галлюцинировал. Такое состояние продолжалось вплоть до конца октября, когда я был приглашен для его лечения на дом. При первом моем посещении больного я нашел его в состоянии сильного угнетения: он постоянно и бесцельно бродил по комнате, на лине его был выражен беспричинный страх, он отвергал всякое исследование, уходил, когда его хотели выслушать, заявляя: «оставьте, мне ничего не надо!». При этом он почти постоянно переходил с места на место, охая и стеная. На всякие расспросы о причине своего страха и стонов стереотипно повторял одну и ту же фразу Ох, что со мною сделали-то!». На самые подробные расспросы он не отвечал ничего определенного о своем состоянии. Можно было только догадываться по боязливому выражению лица больного, что он подвергается слуховым и зрительным галлюцинациям.
Последний факт подтвердился при позднейших моих посещениях больного. Пищу больной принимает сам, но не иначе, как походя; спит плохо; запоры, кишечник вздут, других физических расстройств не имеется. С назначением бромидов, Adonis,a,Codein'a, Laxant, а и ванн угнетение несколько уменьшилось, и хотя в общем больной держал себя попреж-нему, обнаруживая те же явления беспокойного угнетения, охая и стеная, но от него можно было узнать путем многократных расспросов, что он слышит голоса и что на него что-то страшное наговорили, что его детей режут и т. п. При этом сон сделался крепче и больной вообще стал несколько спокойнее. Это относительное улучшение продолжалось и в последующие дни, причем больной стал несколько более доступен при расспросах, но все же давал лишь отрывочные указания относительно своих слуховых галлюцинаций враждебного характера. Голоса незнакомых ему лиц больной слышит, повидимому, постоянно; на него наговаривают, ему угрожают. Он постоянно повторяет «Ох, что со мною будет! Что со мною сделали-то!». Более подробных указаний не дает; хотя и делает попытки рассказать, но затем останавливается, охает и заключает: «Нет, да вы не можете этого понять!». При этом поражает все время отрицательное отношение больного к окружающему. Он заявляет, что исследовать его не нужно, посещать также, упорно просит не прописывать ему лекарств, так как все равно принимать не будет, и т. п. Часто повторяет, что «ему ничего не надо». Тем не менее лекарства больному удается давать после некоторых уговоров; в ванну также садится после некото рого сопротивления.
Вместе с тем у больного поразительным образом нарушено чувство времени. Когда говорили при нем о ваннах, которые длятся от 1/i до х/2 часа, то, вмешивясь в разговор, он упорно утверждает, что ванны длятся секунды. На возражения, что этого не может быть, упорно утверждает, что ванны длятся не более нескольких секунд. Спит он также, по его словам, минутами, хотя, на самом деле, спит хорошо в течение всей ночи. Рубашку ему меняют, по его словам, по 30 раз в день. 6 ноября по возвращении с поездки, во время которой он прокатился от своей квартиры по набережной Фонтанки до Николаевского моста по Неве, что требовало в общем около часу времени при поездке взад и вперед, на вопрос «долго ли ездили?», обращенный к его жене, он тотчас же вмешался, заявив, что ездили всего 2 минуты. На мои возражения он упорно утверждал, что ездили 2—3 минуты, не более! Итак все в том же роде. На вопрос, например, когда я был у него, он отвечает сегодня, недавно, тогда как предыдущее посещение мое было три дня тому назад. В остальном состояние прежнее. Больной вообще крайне несловоохотлив, и от него трудно выпытать подробности его бреда. Из отдельных же заявлений больного можно заключить, что он подвержен галлюцинациям.
18 ноября, будучи спрошен о продолжительности своей поездки, длившейся около часу, заявил, что катался всего минуту, на переспрос ответил: «не более секунды, мне кажется, что я даже и вовсе ие катался!».
382
Но когда ему стали пояснять, где сн был, то он сказал, что он, быть может, даже оставался в своей квартире, а все остальное, о чем ему говорят, он, действительно, видел, но он мог видеть все это раньше. На вопрос, сколько сидел в ванне, отвечает уверенно: «одну минуту». На вопрос, спал ли и как долго, ответил, что спал всего минуту или секунду, что «ночи, повидимому, вовсе и не бывает». На вопрос, сколько он живет на своей квартире, ответил после долгого размышления «5 лет», тогда как в действительности живет 20 лет. Нынешний год определяет правильно, но месяц определяет тот, в который он заболел, т. е. июль, утверждая, что все неверно считают, прибавляя целые месяцы. О событиях прошедших вспоминает, как о событиях недавних. При расспросах часто стонет и охает и галлюцинаторные явления выяснить не удается вследствие несловоохотливости больного по отношению к причине его угнетения.
В декабре месяце галлюцинации несколько обострились. Больной стал утверждать, что детей его зарезали или крошат как капусту, что все делают нарочно для него, что даже фонари на улицах существуют нарочно для него, так как на свете будто бы все должно быть светло, а фонарей никаких не нужно. Говорит о каких-то крокодилах, будто бы даже сам находится внутри одного большого крокодила. Что касается чувства времени, то оно попрежнему изменено в смысле уменьшения протекших промежутков. Больной попрежнему говорит, что сейчас июль месяц. И, несмотря на то, что видит снег из окон своей квартиры, не может разувериться: «По-моему теперь июль, а почему снег, сказать не могу!». Из бредовых идей следует отметить, что, по его мнению, он почти сейчас родился, и, несмотря на всевозможные доводы, он все же утверждает, что ему кажется, будто он только-что родился; он говорит также, что в глазах у него почему-то темно.
В общем состояние больного оставалось без перемены до марта месяца, когда оно стало заметно улучшаться. Мало-помалу угнетение перестало быть столь напряженным, и галлюцинации стали ослабевать.
С половины апреля вместе с развитием более энергичного послабления на низ угнетение больного начало постепенно уменьшаться и он стал постепенно освобождаться и от своего бреда. В течение 2—3 следующих недель бред совершенно исчез и больной в конце концов вполне поправился. По выздоровлении он рассказал о своем болезненном состоянии следующее: слуховых галлюцинаций у больного было сравнительно мало, зато имелись оживленные зрительные иллюзии и галлюцинации. Он видел, например, змей в своей комнате и даже на кровати, видел крокодилов, наконец, ему начало казаться, что он сам находится внутри большого крокодила.
Кроме галлюцинаций, некоторые из бредовых идей больного в основе своей имели иллюзии и перетолковывание впечатлений окружающего. Так, утверждение больного, что жгут его детей, основывалось на том, что детей его уводили из комнаты, а затем в другую дверь вносили дрова для топки печей, что, очевидно, могло быть и в действительности. Точно так же вольной утверждал, что детей рубят, как капусту, очевидно, под влиянием сходственных стуков, слышимых им из хозяйственных комнат. Вместе с этим больному казалось, что люди не умирают, что он только-что родился или точнее переродился, что ему приставили другую голову; что в глазах у него была какая-то темнота и т. п.
Больной ясно вспоминает, что время ему казалось проходящим с необычайной быстротой. Прокатается, например, он около часу, а ему кажется, что он катался всего одну минуту, что он почти не успел выехать, как уже возвратился домой. Между тем, все обстоятельства, сопутствовавшие поездке, он помнит хорошо.
Точно так же и по выздоровлении больной ясно вспоминает, что ему взнны казались минутными, но чем объяснить такое уменьшение времени, ел сказать не может.
383
Из приведенных данных представляется очевидным, что у больного имелось соответственное изменение чувства времени, так как вышеуказанных явлений нельзя объяснить бредовыми идеями больного, что подтверждается и расспросами больного по его выздоровлении.
В других случаях больным, наоборот, сравнительно небольшие промежутки времени кажутся необычайно продолжительными, например, помногу лет. В этом отношении особенно характерным представлялось наблюдение, сделанное мною еще в 1889 г., относящееся к больному, страдавшему алкогольным галлюцинаторным помешательством, которому путешествие от реки Волги до Казани на расстоянии всего не более 7 верст показалось продолжительностью в 100 лет. Вот как описывает по выздоровлении сам больной это свое путешествие:
«Ехали мы, мне так тогда казалось, лет 100 и уехали далеко далеко! Я жену держал за талию и сидел без шляпы все время. Была зима, по мне было ужасно жарко, руки были как в огне. Прошло по меньшей мере столетие! Я заплакал о детях! Где они? И вдруг огни, огни, огни и справа, и слева (т. е. городские огни от уличного освещения при въезде в город) и я приехал во дворец, т. е. в Казанскую окружную лечебницу. Куда девалась жена (это я хорошо помню), я не знаю. Через два года (через сколько времени в действительности, я и теперь не знаю) ко мне пришел д-р И. И. Н. и сказал, что я всего еще в больнице две недели».
Нет надобности доказывать, что и здесь дело идет не о бредовом состоянии, которое объясняло бы заявления больного с преувеличением времени, а о действительном своеобразном нарушении чувства времени совершенно противоположного характера по сравнению с предыдущим случаем.
Аналогичные явления наблюдались мною еще в нескольких случаях, причем также можно было убедиться, что больным самое время казалось значительно более длинным, нежели в нормальном состоянии. Есть основание полагать, что в некоторых случаях бред больных о необычайном долголетии основан на та^ом именно изменении чувства времени, как в только что приведенном случае.
По окончании настоящей работы я имел возможность познакомиться по реферату с докладом д-ра В. В. Воробьева «Случай своеобразного расстройства представлений о времени при прогрессивном параличе», прочитанном им в заседании Общества невропатологов и психиатров при Московском университете от 29ноября 1902 г. Больной д-ра В. В. Воробьева, страдавший типическим прогрессивным параличом при явлениях легкомыслия и эйфории, а по временам угнетения и слезливости и значительного слабоумия мог ориентироваться как в пространстве вообще, так и в окружающей обстановке, но обнаруживал большую дезориентировку относительно времени. Тотчас по поступлении в больницу больной точно определяет число 27 сентября, на другой день определяет 28 сентября, но к вечеру этого дня го-
384
ворит, что еще утро, 11 часов 28 сентября, на следующий день к вечеру больной говорит, что теперь 28 сентября И1/2 часов утра. Таким образом, больной прибавляет в своем счете вместо каждых суток по получасу, впоследствии же еще более уменьшил свой счет и, наконец, на полуночи 28 сентября окончательно остановился в счете времени; смену же дня и ночи объяснял искусственным затемнением; при этом первоначально все события локализовал довольно правильно с расчетом получаса вместо суток, хотя позднее стала обнаруживаться уже неточность в определениях времени. Промежутки в несколько минут больной определял правильно и мог сказать, что обед длится от х/2 до 1 часа времени и т. п. Автор признает вышеуказанное явление не за бред слабоумного, а за особое расстройство памяти времени. При обсуждении этого доклада мнения, однако, разошлись. Некоторые, как В. А. Муратов и И. П. Постовский рассматривают эти заявления больного как нелепый бред, а не как самостоятельное расстройство чувства времени, А. Н. Бернштейн же полагает, что в этом случае дело идет об утрате способности локализовать события во времени.
По И. П. Постовскому, здесь на почве слабоумия могла образоваться бредовая идея о том, что 29 сентября наступить не может, вследствие чего больной при локализации событий во времени прибегает, с одной стороны, к более мелким периодам, чем сутки, т. е. к часам и минутам, а с другой стороны, к большому числу повторного наступления обыденных событий.
Не решая окончательно вопроса о причине нарушенного определения времени у больного паралитика, наблюдаемого доктором В. В. Воробьевым, нельзя не обратить внимания на одно различие между его наблюдением и представленными выше наблюдениями в том отношении, что больной В. В. Воробьева определял небольшие промежутки времени правильно, даже продолжительность того или другого акта определялась больным более или менее точно, тогда как в первом из приведенных мною наблюдений нарушилось определение промежутков времени как больших, так и сравнительно малых в сторону их уменьшения, причем и определение продолжительности отдельных актов оценивалось значительно меньше, чем следует, во втором же наблюдении сравнительно небольшой промежуток времени оценивался больным как сто лет, причем и в том, и в другом случае больные рассказывали об этих нарушениях чувства времени по выздоровлении при нормальной критической оценке своего бывшего состояния. На этом основании есть полное право полагать, что в известных случаях у душевнобольных мы, несомненно, имеем дело с первичным, т. е. необусловленным бредом больных нарушением чувства времени.
25 В. М. Бехтерев
О ЗНАЧЕНИИ ВНИМАНИЯ В ОТНОШЕНИИ ЛОКАЛИЗАЦИИ И РАЗВИТИЯ ГАЛЛЮЦИНАТОРНЫХ
ОБРАЗОВ 1
Уже давно в отдельных случаях психических заболеваний обращала па себя внимание своеобразная локализация галлюцинаторных образов. Особенно поразительно, что больные, в развитии галлюцинаций которых играет очевидное значение психический момент, иногда локализуют свои слуховые галлюцинаторные образы внутри тела, например, нередко внутри своего живота, реже в дыхательном горле и других частях. Из целого ряда имеющихся в нашем распоряжении наблюдений можно было бы привести множество интереснейших примеров, если бы эти факты не признавались в настоящее время более или менее установленными, в которых уже нет более повода сомневаться.
С другой стороны, вообще локализация слуховых галлюцинаций в окружающем пространстве возбуждает много вопросов. Почему одни больные слышат голоса из определенной части пространства, например, из-за стен комнаты, другие —из-под пола или сверху, из-за потолка, с неба и т. п.
В этом отношении прежде всего нужно иметь в виду, что в локализации психологически обусловленных галлюцинаторных образов в большинстве случаев не существует ничего постоянного. Не говоря о том, что часто у больных не имеется определенной более или менее точно фиксированной локализации галлюцинаторных образов, так как они слышат голоса отовсюду, т. е. с разных сторон, но и те больные, которые слышат голоса с одного какого-нибудь пункта окружающего пространства, невсегда указывают одно и то же место появления своих голосов, чаще же заявляют, что их голоса исходят то из одного, то из другого пункта. В других случаях больные, слышавшие голоса с одного пункта, через некоторое время заявляют, что они слышат голоса уже из другого пункта, причем бывает иногда совершенно невозможно уловить причину переселения голосов из одного пункта в другой. Само собой разумеется, что при перемене окружающей обстановки голоса изменяют нередко свою локализацию.
1 Доклад, сделанный в одном из научных собраний клиники душевных и нервных болезней в течьнне 1902 г.
3£6
Так, больные, слышавшие раньше голоса из-под пола, начинают слышать их из-за стен или с потолка и т. п.
Ввиду этого, естественно, возникает вопрос, что же служит причиной того, что слуховые галлюцинации в одном случае локализуются в одном пункте окружающего пространства, в другом случае в другом; иначе говоря, какой психологический процесс определяет ту или другую локализацию слуховых ощущений в окружающем пространстве.
В этом отношении необходимо прежде всего иметь в виду влияние внешних впечатлений на локализацию слуховых обманов. Всякий знает, что нередко слуховые галлюцинации относятся к той части окружающего пространства, откуда слышится шорох или какие-либо иные звуковые впечатления.
Вообще объективные звуковые впечатления, как показывают клинические наблюдения, оказывают огромное влияние на локализацию слуховых обманов, особенно у больных, страдающих слуховой гиперестезией, как нередко случается, например, у галлюцинирующих больных, страдающих хроническим алкоголизмом. Нередко приходится наблюдать, что больной, слышавший раньше голоса из той или другой части окружающего пространства, вместе с тем, как раздастся звон колокола на соседней колокольне, начинает слышать те же самые голоса уже в колокольном звоне; равным образом достаточно бывает больному с слуховыми обманами услышать журчание льющейся или капающей из водопроводного крана воды и он начинает слышать свои голоса в журчаньи текущей воды. Если во время оживленных слуховых галлюцинаций больной услышит стук шагов, то нередко его галлюцинации начинают слышаться в стуке шагов. Иногда даже больные слышат голоса в стуке своих собственных шагов и даже в урчаньи своих кишок, как я убеждался неоднократно.
Исследуя алкоголиков, страдающих слуховыми галлюцинациями, мне удавалось по произволу изменять локализацию этих галлюцинаций. Так, если заставить такого больного прислушиваться к звуку электрического индукционного аппарата, то голоса, раньше слышанные больным с той или другой области окружающего пространства, он нередко начинает слышать в звуке электрической машинки, но достаточно приостановить звук машинки, как тотчас же голоса или временно исчезают или изменяют свою локализацию в пространстве. При возобновлений монотонного звукового раздражения от дребезжащего молоточка индукционного аппарата больной тотчас же снова начинает слышать голоса в звуке электрической машинки. Если последнюю переставить на другое место, то и больной, прислушивающийся к звуку электрической машины, будет слышать свои голоса из того пункта, куда переставлена машина.
Все эти данные не оставляют сомнения в том, что локализация галлюцинаторных образов в известных случаях находится в прямой зависимости от непроизвольного направления внима-
Т>* М7
ния, благодаря чему каждый раз, когда источник звука или, что все равно, какой-либо другой предмет, привлекающий внимание, будет перемещен, то переместятся соответственным образом и галлюцинаторные образы.
В справедливости вышесказанного нетрудно убедиться и на основании одного недавно представившегося мне крайне интересного случая, относящегося к больной, страдавшей около 10 лет слуховыми галлюцинациями вследствие поражения органа слуха. Болезненное состояние больной было уже описано мною в моги статье «О галлюцинаторном психозе, развивающемся при поражении органа слуха» и здесь нет надобности подробно останавливаться на нем, ho заслуживает внимания одна особенность у одной из описанных больных, состоящая в следующем.
Особенностью больной, демонстрированной мной в научном собрании врачей клиники душевных и нервных болезней, является то обстоятельство, что она может вызвать галлюцинации искусственно, производя своей рукой простое прикосновение или легкое почесывание по лбу, по щеке, по губам и по другим частям тела. Эти галлюцинации слышатся больной из той части тела, которую она почесывает, и притом всегда различные по содержанию.
Впервые больная убедилась в появлении у нее галлюцинации при почесывании тех или других частей тела совершенно случайно. Однажды в Оптину пустынь, где жила больная, приехал великий князь К.; больная, очевидно, заинтересовалась его личностью и, когда великий князь стоял в церкви во время богослужения, больная не сводила с него своих глаз и старалась подражать ему во всем. Когда великий князь брал себя за ус и больная прикасалась к своей верхней губе, когда великий князь касался своего подбородка, делала то же и больная, при этом она убедилась, что, кроме голосов, которые она слышала в церкви вообще, она слышала также голос из того места своей кожи, к которой прикасалась. Она подумала, что этим путем великий князь передает ей свои мысли. При этом, производя прикосновение к различным частям тела, например, ко лбу, к щекам, к руке, и прислушиваясь к ним, больная убедилась, что каждый раз слышались те или другие слова из этих областей тела. То же самое больная испытывала и во время пребывания своего в клинике, причем ее нетрудно было заставить вызывать свои галлюцинаторные образы почесыванием тех или других частей тела. При этом оказалось, что больная должна была каждый раз прислушиваться к той области, которую она почесывала, чтобы услышать свои галлюцинации. Очевидно, что и здесь дело сводится к направлению внимания, которое и содействует развитию галлюцинаций.
Что это так в действительности, доказательством может служить тот факт, что больной достаточно прислушаться к какой-либо части пространства, чтобы появилась у нее в этом напрзв-
388
лении слуховая галлюцинация. Так, когда я заставлял больную прислушиваться в направлении определенного угла, то она начинала слушать галлюцинации из этого угла, когда же я заставлял больную прислушиваться к другому углу, тогда она начинала слышать голоса уже из этого угла. Вообще куда бы ни привлечь внимание больной, она оттуда и начинала слышать голоса.
В последнем случае, кроме влияния внимания (в данном случае волевого) на локализацию галлюцинаторных образов, это внимание, очевидно, содействовало и появлению галлюцинаторных образов.
Для проверки положения, устанавливающего зависимость локализации галлюцинаторных образов от непроизвольного направления внимания, я воспользовался одной из бывших под моим наблюдением истеричной больной, у которой нетрудно было вызывать глубокие степени гипноза и внушать в нем самые оживленные слуховые галлюцинаторные образы. Эти галлюцинаторные образы слышались больной с той или другой части окружающего пространства, смотря по тому, где больная воображает по своему разумению, говорящее лицо, но затем достаточно было больную заставить прислушиваться к тому или другому постороннему звуку, как она начинала слышать голоса именно из того места, откуда она слышала звук. Даже если без всякого постороннего звука путем внушения привлекали внимание больной к тому или другому пункту пространства, как больная начинала слышать голоса из той именно области пространства, к которой привлекалось ее внимание.
Даже привлечением внимания к ее животу удалось вызвать слуховые галлюцинации, исходящие из живота больной точ-в-точь, как это наблюдается иногда в патологических случаях у лиц, страдающих хроническими галлюцинациями. В недавнее время опыты такого же рода были проделаны мною в присутствии д-ра Нарбута над другим гипнотиком с влиянием внешних впечатлений не только на слуховые, но и на зрительные и осязательные галлюцинации. Молодой человек Т. был приведен в состояние глубокого гипнотического сна, причем ему были внушены слуховые галлюцинации в виде голоса своего брата, только что к нему пришедшего и говорящего ему о своем житье-бытье. Голос при этом слышится больным прямо спереди, как будто бы его брат стоял впереди его. Затем пускается в ход молоточек индукционного аппарата, поставленного с боку больного, и оказывается, что без всякого постороннего внушения голос своего брата Т. слышит теперь уже в том месте, где шумит молоточек индукционного аппарата, хотя самый шум молоточка им не слышится вовсе. После того как индукционный аппарат незаметно от Т. переставили на другую сторону, и голос стал слышаться им уже из этого нового места, где помещен аппарат; таким образом, сколько бы раз мы не переставляли индукцион-
389
ный аппарат, голос, слышимый Т. в виде галлюцинации, постоянно меняет свое место сообразно местоположению источника зьука, хотя самый звук аппарата и не слышится Т. Очевидно, что источник звука здесь направляет только внимание больного в определенную сторону, чем определяется локализация слуховой галлюцинации.
Достойно внимания, что вместе с прекращением галлюцинации по внушению шум индукционного аппарата начинает вновь слышаться Т., тогда как с появлением внушенной галлюцинации шум индукционного аппарата тотчас же перестает им слышаться. Факт этот, очевидно, следует объяснить поглощением внимания в сторону слуховой галлюцинации.
Совершенно аналогичные результаты были получены и при испытании с помощью камертонов: каждый раз при звучании камертона стой или другой стороны внушенная слуховая галлюцинация изменялась в своей локализации и проицировалась в области источника звука. При этом самый звук камертона не был слышен, пока продолжалась слуховая галлюцинация. С прекращением же последней по внушению звук камертона становился тотчас же слышным для Т.
Совершенно аналогичные явления можно было наблюдать и в отношении зрительных галлюцинаций. Т. в гипнозе была внушена галлюцинация в форме светлой звезды и оказывается, что местоположение этой звезды менялось вместе с тем, как больному был показываем из того или другого места небольшой медный диск, хотя последний Т. и не видел, по его удостоверению. В другой раз ему была внушена зрительная галлюцинация в виде кривляющейся физиономии, причем с той и другой стороны от больного впереди от него было расположено у стен по одной лампочке накаливания. Оказалось, что каждый раз, когда зажигали одну лампочку, галлюцинация, локализовавшаяся первоначально впереди от больного, перемещалась в сторону лампочки, хотя последняя во время галлюцинирования и не замечалась Т., при тушении лампочки галлюцинация опять проицировалась прямо en face от больного, с зажиганием же лампочки, лежащей по другую сторону от больного, снова галлюцинация перемещалась в сторону зажженной лампочки, которая, однако, сама не была доступна восприятию. Опыт этот удавалось проделать бесчисленное число раз и всегда с одним и тем же результатом. Если внимание больного было отвлечено от зажженной лампочки, то она и не влияла на локализацию галлюцинации, но достаточно было зажечь другую лампочку, и тотчас же галлюцинация проицировалась в сторону этой вновь зажженной лампочки. Последняя тем не менее, как уже упоминалось, была невидима для Т. до тех пор, пока, благодаря новому внушению, не исчезала зрительная галлюцинация.
С кожными ощущениями опыт было трудно обставить, но и здесь в общем мы получили сходные результаты. Т. в гипнозе
390
внушали, что его кусает комар. После многократных внушений вызвать этой галлюцинации не удалось; но как только стали прикасаться кончиком бумажки к тылу одной кисти рук как он начал чувствовать укус в этой именно области. Как только больному начали слегка водить той же бумажкой по тыльной поверхности кисти другой руки, как укус стал чувствоваться Т. в месте этого прикосновения на другой конечности.
Заслуживает внимания, что вызывание галлюцинации у Т. в одном органе чувств, например, в органе слуха, обыкновенно не давало возможности вызвать внушением галлюцинации в другом органе чувств, за исключением того случая, когда эта последняя, согласно внушению, должна слиться в то же время с первой в виде одного образа, например, слышание голоса и видение говорящего человека.
Последний факт, повидимому, следует поставить в связь с отвлечением внимания в сторону внушенной галлюцинации, что само по себе не лишено известного научного интереса.
Все эти наблюдения не оставляют сомнения в том, что та или иная локализация галлюцинаторных образов, как слуховых, так, очевидно, и иных, объясняется привлечением внимания больных к той или другой части окружающего пространства в силу каких-либо объективных явлений, наблюдающихся в данной области. Очевидно также, что и локализация галлюцинаторных явлений внутри своего тела объясняется привлечением внимания галлюцинирующих больных к данной области.
В этом отношении нетрудно привести много примеров, где больные локализовали свои слуховые галлюцинации внутри своего тела в зависимости от тех или других ощущений. Так, в одном случае дело шло о женщине, слышавшей голоса изнутри сво#го тела, при вскрытии уже у этой больной оказалось раковое новообразование в кишечнике. Другой больной, страдавший хроническим катарром кишечника, слышал по временам голос, исходивший из области живота, и можно было точно выяснить, что этот голос он слышал в то именно время, когда появлялось у него усиленное урчание в животе. Другой больной слышал голос, исходящий из предсердечной области, который он признавал за голос святого духа, поселившегося внутри тела, и в этом случае можно было проследить появление этого голоса вместе с возникновением чувства необычайной легкости в предсердечной области, которую больной и аллегоризировал в виде присутствия святого духа.
Особенно поучительным в этом отношении мне представляется наблюдение, сделанное И. П. Мержеевским по отношению к больному, страдавшему слуховыми галлюцинациями, которые он слышал, как обыкновенно, извне, но с тех пор, как он заболел перитонитом, те же голоса он начал уже слышать внутри своего живота. Это наблюдение, очевидно, прямо говорит в пользу высказанного нами положения о значении непроизвольного вни-
391
мания в отношении локализации галлюцинаторных образов. Очевидно, что с развитием перитонита внимание больного, помимо его воли, направлялось на область живота, в котором испы-тывались болезненные ощущения, в силу чего и галлюцинации, раньше слышавшиеся больным извне, стали слышаться внутри своего живота.
Даже не одни только патологические чувственные образы, в известных случаях, благодаря направлению непроизвольного внимания, относятся к тем или другим частям своего тела. Есть основание полагать, что и патологические аффективные состояния и общие ощущения могут при известных условиях, благодаря опять-таки направлению внимания, относиться к тем или другим областям, где их локализация представляется явно ненормальною.
Этим, например, объясняются заявления больных, что они испытывают «страх в мочевом пузыре» или что у них «тошнит левую ногу». При этом более внимательные расспросы показывают, что дело идет в этом случае не о фигуральном лишь выражении, а о таких состояниях, которых больные иначе и выразить не могут.
Этим же направлением непроизвольного внимания можно объяснить и прямой переход в известных случаях галлюцинаций в иллюзии и, наоборот, переход психопатологических иллюзий в галлюцинации.
Мы уже выше упоминали о том, что больные, слышащие голоса из какой-либо части пространства, нередко вместе с появлением постороннего звука, например, колокольного звона, начинают слышать голоса в этом звуке, как будто бы те же самые голоса стали, по выражению одного из моих больных, «вызваниваться» в колокольном звоне. Между тем с прекращением колокольного звона голоса попрежнему слышались больным из окружающего пространства. Точно так же вместе с появлением шума текущей воды голоса больного, ранее слышавшиеся из какой-либо области окружающего пространства, начинают слышаться в журчании воды и т. п. Наблюдаемый прямой переход галлюцинаций в иллюзии объясняется, очевидно, невольным привлечением внимания к тем или другим звуковым впечатлениям, вследствие чего галлюцинации и сливаются с последними, превращаясь в иллюзии.
Равным образом и локализация галлюцинаторных образов в окружающем пространстве стоит в известном отношении к привлечению внимания теми или другими внешними поводами. Поэтому в одной обстановке больные слышат свои голоса в одной части пространства, тогда как в другой обстановке те же больные слышат свои голоса уже в другой о'бласти.пространства, смотря по тому, куда по преимуществу привлекается их внимание.
Выше мы привели наблюдение, где до очевидности ясно обманы чувств вызывались под влиянием напряжения в известном
392
направлении. Есть и другие примеры из психопатологии, где можно с положительностью говорить об усилении и появлении галлюцинаторных образов под влиянием напряженного внимания. Особенно нередко у алкоголиков удается вызвать развитие зрительных или слуховых галлюцинаций простым возбуждением вниманият.
Чтобы выяснить эту сторону дела, нельзя не указать на значение внимания в апперцепции вообще и на развитие перцепиру-емых образов.
Известно, что ввимание представляет собой такой процесс, при посредстве которого наши внутренние явления становятся более яркими. Благодаря вниманию то или другое ощущение или идея становятся яснее или, как иногда говорят, выдвигается в поле ясного сознания. Как известно, особенно Вундт выдвинул в последнее время значение внимания в процессе апперцепции, хотя роль внимания в этом отношении была указана еще Лейбницем.
Не подлежит, однако, сомнению, что внимание не только усиливает внутренние явления, но оно в состоянии и возбудить те или другие внутренние образы. Это может быть доказано экспериментальным путем при работах с гипповским хроноскопом. Всем работавшим с этим аппаратом известно, что когда требуется отмечать простую реакцию, то нередко дается сигнал во время самого звука или даже ранее его появления, несмотря на то, что лицо, над которым производят опыт, убеждено, что отметки сделаны верно.
Если мы возьмем маятник Вундта и установим звонок так, чтобы звон раздавался на определенном пункте положения маятника, то при обращении нашего внимания не на самый звук, а на любой пункт полукруга с делениями, по которому двигается конец маятника, оказывается, что мы слышим звонок каждый раз в том пункте, на который мы обращаем внимание, а не на том месте, на котором установлен звонок прибора.
С другой стороны, если мы возьмем карманные часы, ход которых может быть приостанавливаем по желанию исследователя и незаметно для испытуемого лица, то, отведя часы на такое расстояние, чтобы бой их был едва слышен, мы можем привлечением внимания испытуемого субъекта заставить слышать бой часов в то время, когда его наверно уже нет.
Все эти факты, очевидно, говорят в пользу того, что внимание является таким актом, который способен возбудить внутренние образы там, где их нет в действительности. В повседневной жиз;
В. Бехтерев, Об искусственном вызывании обманов чувств у алкоголиков, страдающих галлюцинаторными формами помешательства. Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 2, стр. 101—104.
393
ни мы встречаем подтверждение этого везде и всюду. Когда внимание привлечено в каком-либо направлении, то всякий знает, чго мы получаем нередко действительные иллюзии, т. е. создаем образы, не соответствующие действительному восприятию. Очевидно, что и в патологических случаях непроизвольное внимание может оказывать возбуждающее влияние на появление галлюцинаторных образов, как это мы и наблюдали в приведенных примерах.
ПРИЗНАНИЕ ДЕЙСТВИТЕЛЬНОСТИ
ЗА ГАЛЛЮЦИНАТОРНОЕ ПЕРЕЖИВАНИЕ КАК ОДНО
ИЗ ПРОЯВЛЕНИЙ ПСИХИЧЕСКОГО РАССТРОЙСТВА
В числе многочисленных симптомов психических заболеваний заслуживает внимания то своеобразное состояние, когда больные окружающую действительность признают не за действительность, а за нечто искусственное, подставное или за продукт своих галлюцинаций или сновидения, или, наконец, за продукт собственной фантазии. Больные в этом случае все воспринимают совершенно правильно, как оно и есть в действительности, но не считают окружающее за действительно переживаемое, а за нечто фантастическое, представляющее лишь их собственные галлюцинации или сновидения, или же признают за намеренно подставленное другими. Представим раньше всего примеры подобного рода состояний.
В одном из моих наблюдений дело идет о девице в возрасте около 42 лет, надзирательнице одного из больничных учреждений Петербурга.
Больная имеет семейное наследственное расположение к нервным болезням и сама с давних пор отличалась нервностью, доходившей до приступов малой истерии. В остальном больная всегда была здоровой, но за последние годы у нее стал развиваться фиброид матки, достигший величины детской головки. В течение последних двух месяцев месячные отсутствуют совершенно.
При исследовании физической сферы в сентябре 1905 г., кроме этих изменений со стороны половой сферы, никаких других изменений не обнаружено. С физической стороны можно лишь отметить умеренное повышение сухожильных рефлексов и ослабление глоточного рефлекса. В последнее время, за период 1905 г. больная пережила несколько тягостных для нее моментов нравственного характера, под влиянием которых со стороны психической сферы с конца лета 1905 г. у нее начали проявляться уклонения, выражавшиеся в том, что она стала относиться к к прошлым событиям и ко всему окружающему не как к действительности, а как к чему-то искусственному или подставному, или составляющему продукт ее галлюцинаций. Больная, например, в течение летнего времени участвовала в похоронах своих сослуживцев и хотя во время этих похорон она все признавала их за действительность, но затем ей стало казаться, что это была лишь галлюцинация. Равным образом ей казалось, что вся окружающая обстановка и люди, с которыми она беседует, не являются действительными, а оказываются чем-то подставным, нереальным. Подобное состояние у больной длллось в течение более месяца Никаких галлюцинаций, никаких иллюзий больная в это время не имела.
Вышеуказанное состояние у больной длилось всего несколько недель и затем совершенно исчезло. Но в самое последнее время, спустя почти год после вышеуказанного состояния, больная, как я узнал, заболела вновь психическим расстройством, на этот раз более тяжелым, потребовавшим помещения больной в специальное заведение.
В этом примере мы имели психическое расстройство, не достигшее вполне выраженного психоза и обусловленное, по всей вероятности, нарушением кровообращения, связанным с подавлением menstrua в климактерическом периоде у субъекта, предрасположенного к психическим расстройствам. Приведу еще другой пример.
Больная К-, 24 лет, девица; мать се нервная; бабушка с головными болями; отец с состоянием угнетения, умер от последствия покушения на самоубийство. Брат деда умер психически больным. Сама больная с детства нервная, в последнее время стала уставать па службе и нервность ее усилилась. С год назад ей стало казаться временами, что виденное ею ранее представляется либо сновидением, либо нереальным, а пережитым, как продукт своей фантазии или грез, и в то же время кругом себя она видит не действительность, а все окружающее ей только кажется или оно есть ни что иное, как сновидение. Недавно, например, больная ждала с нетерпением к себе знакомого; но, когда он приехал и беседовал с нею, ей все казалось, что это не действительность, а кажущееся явление, своего рода галлюцинации. Что бы больная ни видала вокруг себя, ей кажется, что это не действительность, я нечто нереальное, кажущееся. Если ей что-нибудь говорят, она также не признает это за действительность, а за мнимые голоса мнимых людей. Она иногда сама хочет себя убедить в том, что все окружающее есть действительность, но это ей не удается. Если ее уверяют, что все ее окружающее на самом деле существует, то она тем не менее этому не верит. Также и относительно прошедших событий больной кажется, что бывшее с ней представляло ее галлюцинации или ее фантазию, или, быть может, было во сне. Никаких обманов чувств больная не испытывает. Память и ассоциации правильны. Вообще в остальном чувствует себя здоровой. Сон не всегда спокойный, иногда испытывает продолжение сновидений некоторое время после пробуждения. Больная сама записала о своем состоянии следующее:
«Сны мои были прежде продолжением действительности или продуктом моих грез иаяву(яочень много мечтала, очень любила это занятие и предполагала долго жить среди выдуманных мною сказок, осуществления которых я не ждала и в которые уходила от действительности); теперь это масса спутавшихся образов и фраз из повседневной жизни; благодаря вечному туману в голове, мне трудно разобраться, какое слово> или явление принадлежит сну и какое — действительности (иногда это нужно), тем более, что я очень скоро забываю даже то, что было вчера (прежде этого не было). Два или три раза в последнее время (в минуты сильной тоски, предчувствия несчастья или сильного потрясения) у меня являлось сомнение в реальности окружающего меня; мне казалось, что все, что делается кругом,— результат напряженного воображения, что, может быть, все совершенно не так и нет того, что сейчас у меня перед, глазами: я не сижу перед столом, я не вижу своего собеседника (т. е. этого нет на самом деле), я где-то в другом месте, но мозг мой выдумал все то, что представляется моим глазам и слуху; мне казалось, что я,, может быть, уже сумасшедшая и галлюцинирую, и это было ужасно.
Я в эти минуты не отрицала реального существования и своих родных, и обстановки, но ощущение было вроде того, как будто я, лежа в своей комнате, выдумала, что я иду на службу, что со мной разговаривают, что приезжают те, кого я жду, а ничего этого сейчас нет. Я разуверяла себя тем, что все совершающееся кругом уже очень ярко и последо-
396
вательно, как не бывает во сне или в бреду, а сомнение подсказывало, что у сумасшедших галлюцинации дают полную иллюзию действительности».
Кроме всех вышеуказанных явлений, у больной имеются ясные неврастенические симптомы. Жалобы ее сводятся на усталое подавленное состояние, отсутствие аппетита, прерывистый, неспокойный сон, постоянное чувство беспокойства, туман в голове, вызывающий смутность мыслей, кажущееся ослабление памяти и интереса к событиям, постоянный шум в голове, неспособность что-либо написать без насильственного возбуждения, тоски, беспокойство, наряду с усталостью и тупостью, и недовольство окружающим.
В последнем случае дело идет, таким образом, о неврастеническом и частью, может быть, истерическом состоянии, в котором вышеуказанный симптом выражен в резкой степени. Но одинаковые явления наблюдаются и в тяжелых психозах, особенно острых, например, при галлюцинаторной спутанности, и притом в общем далеко нередко, но здесь они часто затемняются рядом других симптомов и потому не всегда легко выделяются из общей картины заболевания. Тем не менее, несомненно, что эти явления нередко играют важную роль в развитии бредовых идей, наряду с действительными обманами чувств.
Во всех этих и подобных им случаях дело, очевидно, идет не о нарушении процесса восприятия, которое у наших больных оставалось вполне правильным и совершенно ненарушенным, а в нарушении более сложного психического процесса — процесса узнавания. Дело в том, что больные, вполне правильно воспринимая окружающее, не могли лишь его признать за действительность, несмотря на то, что их процесс суждения в то же время не 1 представлял существенных уклонений от нормы.
Дело идет, таким образом, о настоящих обманах узнавания, которые могут наблюдаться в известных случаях более или менее изолированно, в других же случаях наблюдаются наряду с обманами чувств и другими нарушениями психической сферы.
Следует заметить, что в отдельных случаях больные не признают и своих сновидений за действительные сновидения, а рассматривают их как искусственно вызванные явления. Так, один из наших больных, не обнаруживавший в бодрственном состоянии никаких иллюзий и галлюцинаций, испытывал всевозможные и крайне оживленные сновидения. В этих сновидениях он видел цинические картины, в которых проделывались будто бы «невозможные вещи» с его женой. Все эти сновидения он признавал за искусственнно показываемые ему картины, resp. галлюцинации, вызванные путем гипноза какими-то злоумышленниками, и мучился мыслями о возможности действительных проделок такого рода с его женой.
Рассмотренные состояния представляют собой совершенно своеобразный симптом, который можно сблизить с теми патологическими явлениями, которые известны в литературе под названием парамнезий.
397
Литература, относящаяся к различным формам парамнезий, начиная с 1872 г., представлена целым рядом авторов (Зандер, Янсен, Буарак и Горвиц, Шюле, Пик, Аниэль, Форель, Рибо, Крепелин, Фуилье, Дюга, Саланд, Арно, Леруа, Тололанска и др.). Из позднейших работ, относящихся к этому предмету, можно указать на статью профессора А. Пика 1, затем на статью М. Киндберга2 и др.
В последней работе приведена, между прочим, и литература, относящаяся к парамнезии, а также рассмотрены взгляды авторов на происхождение парамнезии. В отношении последней мы, к сожалению, до сих пор не имеем еще удовлетворительного объяснения и можно сказать, что существует столько взглядов, сколько и авторов, писавших по данному предмету. По Кинд-бергу, при утрате чувства реальности дело идет об утрате ощущения мозговой работы или усилия при опознавании предметов, вследствие чего и нарушается оценка видимого. Больным реальное кажется в виде образов, сна, им все кажется странным, начиная со своих ощущений, они утрачивают чувство личности и чувствуют себя как бы автоматами.
Тот вид парамнезии, который мы описали выше, однако, не может быть объяснен с точки зрения Киндберга, так как, кроме ощущения, сейчас видимого в форме галлюцинаторных образов или сновидений, две наши больные свои воспоминания о прошлом признавали не забытое в действительности, а за пережитое в галлюцинациях, хотя во время самого переживания они этого ощущения не испытывали.
Равным образом и в третьем нашем случае больной, считавший пережитые им сновидения за галлюцинации, вызванные ' в нем путем гипноза, производил неправильную оценку своих состояний после уже совершившегося их переживания, следовательно, в своем воспоминании. Очевидно, что чувство мозговой работы или усилия в наших случаях при самом опознавании не представлялось нарушенным, так как в момент переживания парамнезии не имелось, кроме нашего второго случая, где одновременно существовала и обычная форма парамнезии, выражающаяся неузнаванием окружающей действительности. Отсюда очевидно, что чувство усилия в происхождении этого явления в данных условиях не могло играть какой-либо роли.
Поэтому правильнее смотреть на приведенные нами своеобразные расстройства, как на ошибки узнавания, зависящие не от нарушения процесса восприятия или по крайней мере не его одного, но от нарушения процесса воспроизведения.
1 A. Pick, Patbologie des Bekanntbeitsgefubls und Bekannt-he^ibqualif3(, Neurol. Zbl., 1903, N. 1.
2M Kind berg. Les sentiments du dejS vu et I* illusion de la fausse reconnaissance, Revue de psychiatrie et de psychologie experimentale, 1903, N. 4, и др. •
О ГАЛЛЮЦИНАТОРНЫХ ВОСПОМИНАНИЯХ
Под названием «галлюцинаторные воспоминания» могут быть-понимаемы те своеобразные болезненные состояния, когда больными в виде галлюцинаций воспроизводятся бывшие с ними происшествия или виденное и слышанное ими. Это не что иное, как в настоящем смысле слова галлюцинаторное воспроизведение прошлых, пережитых ранее представлений. Но прежде обсуждения этих своеобразных явлений приведем здесь несколько относящихся сюда наблюдений.
Больная, пожилая девица, без тяжкого наследственного предрасположения, жившая при монастыре, заболевает ушами после инфлюэнцы, причем у нее в конце концов развилось омертвение обеих барабанных перепонок и она получила резкое ослабление слуха, продолжающееся и по настоящее время. При монастыре больную определили в чеканную мастерскую, где она должна была заниматься с девочками. Под влиянием сильного нервного напряжения на почве поражения ее ушей здесь с ней развиваются иллюзии и галлюцинации и она заболевает галлюцинаторным психозом на почве поражения органов слуха. Она начинает слышать-при стуке молоточком в чеканной мастерской слова, а затем ей в стуке стали слышаться целые фразы. Больная стала также слышать свое имя и отчество, произнесенные присутствующими при работе монахинями. Тут же больная была поражена тем обстоятельством, что она услыхала весь разговор, бывший год назад между ней, девочками и надзирательницей в приюте, где она ранее занималась с девочками рукоделием. Дело-происходило так.
Девочка Е. Я. сильно нагрубила, вследствие чего наша больная, тогда бывшая здоровой, отправила девочку из своей рукодельной в младшее отделение; потом надзирательница позвала нашу больную и начала упрашивать снова взять к себе провинившуюся девочку, бывшую ее приемной дочерью, так как девочка сил ьно плакала, двое суток не спала,. не пила и не ела. Также и другие лети долго и настойчиво, более двух дней, упрашивали больную простить девочку и снова взять ее к себе в рукодельную. Больная в конце концов заставила девочку просить прощение за свои грубости и снова взяла ее к себе в рукодельную И вот весь этот разговор и упрявпшиание девочек, уговаривание простить со стороны надзирательницы, плач и мольбы провинившейся девочки и, наконец, слова самой больной со всеми мельчайшими подробностями были воспроизведены в галлюцинаторных слуховых образах с такой точностью, «как будто бы,— по словам больной,— предо мной сейчас все происходило». Больная всем происшедшим была сильно поражена, рассказала обо всем игуменье, которая признала это «искушением» и тем вызвала недовольство к себе у больной.
В другой раз больная услыхала голос своего давнего знакомого П.; он начал рассказывать ей всю свою жизнь, начиная с молодости, а затем
ЗУУ
он начал рассказывать свою позднейшую обыденную жизнь и свои отношения к другим, что больной вообще было хорошо известно. Здесь повторялось все, что больная знала об этом господине с такой точностью, что на нее напал сильный страх. Больная эта, страдающая и по сие время хроническими галлюцинациями слуха, передает и много других галлюцинаций, которые являются настоящим переживанием прошлого.
Другой больной, алкоголик, страдающий алкогольным галлюцинаторным психозом, рассказывая о своих галлюцинациях, между прочим, записал следующее: «Утром, идя на службу в Сенат, ко мне подошла какая-то женщина и попросила на бедность, но так как у меня не было денег, я отказал ей и хотя ее уже не было, но я все до дверей самого Сената слышал ясно и отчетливо ее голос за собой: «господин студент, помогите мне», даже чувствовал, как она за плечо останавливает меня несколько раз, молящим голосом прося помочь ей».
«На службе в Сенате в этот день мне не было легче, голос женщины попрежнему продолжал назойливо обращаться ко мне, а особенно мучили меня страдания прошлой ночи. Стоило мне подумать, как говорила ночыодевочка: «дядя, зарежем его» (это была галлюцинация) и я сейчас же вместо голоса женщины слышал эти слова ясно; или мысленно припомню какую-нибудь фразу, хотя бы, например, сестры своей, которая живет в Томске, и уже слышу: ту же самую фразу кто-то громко произносит около меня точно такой же интонацией и точно таким же голосом».
В этом примере галлюцинаторное воспроизведение пережитых событий выступает не менее резко, нежели в предыдущем случае.
Само собою разумеется, что подобного же рода «галлюцинаторные воспоминания» наблюдаются и в форме зрительных образов.
Так, один больной, страдавший обыкновенными галлюцинациями, рассказывал, что, переехав на большое расстояние от города, в одном из ресторанов которого он кутил, он вновь увидел всю ту обстановку ресторана, где он сидел за столом вместе со своими товарищами, причем видел в числе других товарищей и самого себя. По словам больного, «все представилось ему так, как было в действительности».
Подобные же зрительные галлюцинации в виде точного воспроизведения виденного и пережитого случались с больным и в другом роде.
Другой из моих больных, страдавших галлюцинаторным помешательством, заявлял, что ночью после того, как он проснется, он начинает слышать с мельчайшими подробностями тот самый разговор окружающих лиц, который он слышал днем. Галлюцинация в этом случае достигала полной объективности, так как больному казалось, как будто бы ему кто-нибудь извне шептал этот разговор.
В эпилепсии мне случалось неоднократно наблюдать, что галлюцинации, воспроизводящиедействительнобывшие восприятия, являются в виде особого предвестника приступа. В этих случаях каждый раз падучный приступ начинался повторным воспроизведением в форме галлюцинации воспринятого ранее слухового впечатления. Таким образом, если, например, с больным начинается приступ в то время, когда последнее достигшее до его уха слово из разговора, который он вел с окружающими лицами, было «человек», то это слово «человек» затем воспроизво-
400
дилось в виде галлюцинации много раз и такое воспроизведение продолжалось до тех пор, пока он совершенно не утрачивал сознания. В других случаях перед каждым приступом стереотипно появляется одна и та же галлюцинация, являющаяся повторением бывшего восприятия.
У одной женщины 42 лет обнаружились обморочные припадки 2 раза. Кроме этого, никаких указаний на падучую не имеется; но у нее обнаруживаются крайне оживленные галлюцинации, которые развиваются следующим образом: больная с самого начала «задумается» и за тем подвергается видениям, а иногда и голосам. Случалось, например, что ей покажется посторонний человек, которого она принимает за действительность, крайне пугается этого, а затем как бы очнется и все проходит, причем больная помнит о всем происшедшем, но как будто виденное ею было во сне. Однажды она услыхала голос, приказывающий ей итги на чердак и, повинуясь ему, она, действительно, поднялась на чердак, где увидела рыжего большого человека, предлагавшего ей веревку. Она испугалась и очнулась. В другие разы она тоже начала подниматься на чердак, но очнулась уже тогда, когда хватилась за скобку. У той же больной нередко оживляются в форме галлюцинаций и прошлые воспоминания. Однажды, например, ей представилась вся обстановка ее собственных родов, т. е. она видела перед собой в галлюцинаторных образах всех лиц, бывших при ее родах, а также всю бывшую при этом обстановку и т. п.
К той же категории явлений, повидимому, относятся и наблюдения, в которых галлюцинации, развивавшиеся перед приступом падучей, являлись воспроизведением сновидений, бывших с больным в предшествующую ночь. Подобные случаи, по моим наблюдениям, вообще не составляют редкости. В одном из моих случаев галлюцинации, олицетворявшие бывшие сновидения, представлялись крайне живыми и сложными. Больной, например, во сне видит себя в поле за посевом (он новобранец из крестьян), видит галок, собирающих в поле зерна, слышит их крики и т. п. И это сновидение во всей своей полноте и с точностью воспроизводится в виде галлюцинаций на следующий день перед наступлением падучнаго приступа.
В другом случае больной, молодой человек, страдал издавна головными болями. В последнее время стали присоединяться симптомы неврастении; между прочим, он стал спать тревожно и испытывать массу сновидений, которых он по пробуждении припомнить не в состоянии. Но в течение дня, когда у него развивается головная боль, он начинает подвергаться грезам, в которых он узнает бывшие с ним сновидения. Эти грезы отличаются бессвязностью, состоят из линий, лиц и т. п. Хотя по пробуждении от сна он не может установить ни одного сновидения, но теперь он узнает сразу, что это те сновидения, которые с ним происходили в течение предшествующей ночи. Явления эти продолжаются уже в течение около года. При этом у больного с объективной стороны не было обнаружено ничего существенного.
26 в. М. Бехтерев
401
В другом случае дело идет о больном, страдавшем органическим поражением мозга на почве бывшего lues'а. Объективно у него обнаруживается опускание левого верхнего века, то усиливающееся, то ослабевающее, по временам двоение в глазах и ограничение поля зрения. Из субъективных жалоб больного обращают на себя внимание головные боли в верхней части головы и за левым виском, которые временами достигают необычайной силы и даже однажды сопровождались рвотой.
Гольной, между прочим, рассказывает, что достаточно ему побывать в бане при повышенной окружающей температуре, особенно если он обливается очень теплой водой, как с ним наступает состояние отупения, при котором он видит то, что ему представлялось во сне, и он начинает в настоящем смысле слова галлюцинировать. В это время он начинает суетиться и болтать, что его вообще облегчает, и он быстро приходит в себя. Во время самого видения больной заявляет, что видит то самое, что видел во сне под утро в предшествующую ночь, хотя подробности он помнит смутно, неясно.
В вышеуказанных случаях мы имеем, таким образом, дело с настоящим галлюцинаторным воспроизведением в патологическом состоянии пережитых однажды умственных образов в виде внешнего восприятия или в виде сновидения. Прототипом этого своеобразного патологического явления, что касается по крайней мере галлюцинаторного воспроизведения бывшей действительности, на мой взгляд, может служить воспроизведение действительного восприятия в сонных грезах. Известно, что некоторые лица то, что они видят и слышат днем, переживают ночью во время сна в виде сонных грез или сновидений.
В некоторых случаях это явление обнаруживается в поразительно резкой степени, как показывает, например, нижеследующий случай.
Больная 68 лет, замужняя, без наследственного расположения, имеет 5 человек детей; при последней беременности получила парез обеих правых конечностей, который с течением времени еще несколько усилился. Объективное исследование показало, кроме правостороннего пареза руки и ноги, еще и легкий перез правого facialis и небольшое отклонение языка вправо, а кроме того, крайне резкое повышение сухожильных рефлексов на правой стороне.
При расспросах выяснилось, что больная уже с детства отличается той особенностью, что во сне она переживает все заинтересовавшее ее в течение дня с совершенной точностью. Таким образом, например, побывав в опере, она каждый раз переживает во сне всю оперу от начала до конца в правильной последовательности со всеми участвующими лицами. Это до такой степени для нее представляется обычным явлением, что супруг ее, шутя, замечает, что ей очень выгодно ходить в театр, так как за одни н те же деньги она прослушивает оперу дважды. Равным образом и все, что больную сильно заинтересует или поразит в течение дня, она переживает непременно во сне. Обыкновенно спит она в таких случаях не очень спокойно и часто пробуждается, причем может рассказать все виденное во сне со всеми подробностями. В остальном сон совершенно нормален. Надо при этом заметить, что больная отличается особеи-
402
ной впечатлительностью и, например, возвращаясь поел*; оперы, она сосредоточивается на своих впечатлениях в такой степени, что раздражается, если ее отвлекают какими-либо вопросами.
Хотя приведенный случай и представляется несколько необычайным по крайней живости переживания действительности в форме сонных грез, но известно, что подобное переживание не в столь резкой степени не составляет редкости. Во всяком случае оно может быть поставлено в известную аналогию с переживанием действительности и самих сновидений в галлюцинаторных образах. Сходство обоих явлений увеличивается еще тем, что в отдельных случаях, как это было, например, в последнем из приводимых случаев, состояние галлюцинаций сопровождалось состоянием помрачения и отупения с явлениями амнезии.
Что же касается эпилепсии, в которой также наблюдается галлюцинаторное переживание восприятий, то в наблюдаемых мною случаях оно происходило часто в виде ауры, следовательно, в состоянии уже начинающегося приступа и потому в этих случаях галлюцинаторное переживание может быть также уподоблено сновидному состоянию с переживанием действительности во сне.
Известно, что и в течение больших истерических приступов содержание галлюцинаций нередко составляют бывшие, пережитые ранее больными события.
Из приведенных примеров очевидно, что «галлюцинаторные воспоминания» могут наблюдаться при разнообразных психических расстройствах, как, например, при истерии, эпилепсии, при хроническом алкоголизме, при галлюцинаторном помешательстве и даже при органическом поражении мозга.
Без всякого сомнения такого рода галлюцинаторные переживания прошлого не лишены известного значения с точки зрения патогенеза галлюцинаций, в силу чего они и заслуживают изучения.
26*
ОБ ОТНОШЕНИИ МЕЖДУ ПСИХИЧЕСКИМИ И НЕРВНЫМИ БОЛЕЗНЯМИ
В группу психических, или душевных, болезней входят все те болезненные процессы, в которых преобладающими явлениями служат расстройства со стороны психической, resp. нервно-психической деятельности, иначе говоря, расстройства в области тех отправлений, благодаря которым устанавливается отношение личности к окружающему миру. Из этого определения нетрудно усмотреть, что психические болезни ничуть не выражаются исключительно одними только изменениями в сфере психики (resp. нейро-психики), но что они могут сопровождаться также и другими расстройствами и, между прочим, изменениями со стороны тех или других проводящих функций нервной системы. И действительно, знакомясь ближе с различными формами психических, или душевных, болезней и, в частности, с теми расстройствами, которыми они выражаются, нетрудно убедиться, что в сущности нет или почти нет ни одной психической болезни, в которой рядом с расстройствами психической сферы не обнаруживались бы те или другие нарушения со стороны проводящих функций нервной системы и органов тела, подчиненных деятельности нервной системы, хотя бы и слабо выраженные.
Факт этот становится вполне понятным, если мы примем во внимание, что и те, и другие болезни, т. е. так называемые психические и собственно нервные, относятся к поражению одной и той же нервной системы.
Современная медицина, вопреки недавнему прошлому, даже объединяет между собой психические и нервные болезни, так как выяснилось, что психическая деятельность находится в полной и безусловной зависимости от деятельности высших корковых центров нервной системы и их взаимных связей между собой, в то время как поражение других центров и проводников нервной системы обусловливает разнообразные нервные болезни.
Независимо от этой анатомо-физиологической точки зрения, устанавливающей, так сказать, природную связь между психическими и нервными болезнями, имеются между теми и другими несомненное родство и в этиологическом отношении. Особенно доказательные факты в указанном отношении можно почерпнуть
404
в наследственности. Как известно, в настоящее время точно установлено, что в семьях, отягченных неблагоприятной или патологической наследственностью, мы встречаем, наряду с психическими, также и разнообразные нервные болезни.
С другой стороны, известно, что от лиц, страдавших психическими болезнями, происходит потомство, отличающееся не только теми или другими уклонениями психической сферы, но и поражаемое нервными болезнями, особенно общими неврозами, и, наоборот, от лиц, страдавших нервными болезнями, особенно общими неврозами, обычно происходит потомство, в котором может быть отмечено то или другое количество психопатов.
Словом, между психическими и нервными болезнями существует самое тесное родство, какое только можно себе представить в наследственном отношении.
Равным образом и важнейшие этиологические моменты, помимо наследственности, являются общими для развития психических и нервных болезней. Возьмем для примера травму, нравственный шок, сифилис, разнообразные острые инфекции и другие отравления, а также различные самоотравления. Как известно, все эти условия способствуют в той или иной степени развитию как психических, так и нервных заболеваний. Нередко даже результатом воздействия вышеуказанных причин являются болезни, в которых психические и нервные симптомы представляются как бы смешанными между собою.
Таковы травматические нейропсихозы, нейриты с психическими расстройствами, сифилитические поражения мозга с психическими дефектами, алкогольный ложный паралич и т. п.
Наконец, нельзя не отметить здесь того обстоятельства, что при самых разнообразных психических болезнях при исследовании открываются те или другие нервные расстройства. Это положение справедливо не только по отношению к так называемым органическим психозам, но и к группе истерических, эпилептических, неврастенических, а также в известной мере и к группе дегенеративных и других психозов.
С другой стороны, при многих нервных болезнях, особенно при так называемых общих неврозах, а также при разнообразных поражениях головного мозга (опухоли, размягчения, кровоизлияния, склерозы и пр.) могут быть открыты те или другие нарушения психической сферы.
Далее родство между психическими и нервными болезнями совершенно ясно устанавливается тем фактом, что в известных случаях нервные болезни осложняются психическими расстройствами. Здесь должны быть упомянуты психозы, развивающиеся на почве табеса, гентингтоновской хореи, геморрагического энцефалита и пр. С другой стороны, имеются болезненные психические формы, в течение которых развиваются те или другие нервные поражения. Сюда относятся артериосклероти-ческие психозы, сифилитические психозы и т. п.
405
Мы знаем затем целый ряд болезненных форм, в которых психические и нервные расстройства идут рука об руку в такой степени, что трудно даже решить, куда правильнее относить эти расстройства — к нервным или психическим болезням. Сюда относятся некоторые общие или большие неврозы, как истерия, миастения и неврастения, отчасти эпилепсия, некоторые фамильные нервные болезни, сопровождающиеся слабоумием и т. п.
Как известно, по отношгнию к истерии в последнее время, благодаря трудам Шарко, Мебиуса, Шгрюмпеля, Жане, Реймона и др., все более и более устанавливается взгляд, что несмотря на ряд симптомов, сближающих эту форму болезни с чисто нервными поражениями, она должна быть рассматриваема как психическая болезнь, не потому только, что здесь обнаруживаются чисто психические симптомы, как своеобразные особенности характера, склонность к развитию галлюцинаций, навязчивых идгй и т. п., но и потому, что целый ряд симптомов при истерии, на перзый раз кажущихся нервными, как параличи, судороги, контрактуры и пр., могут быть вызываемы и устраняемы путем так называемого психического воздействия. В самом деле, если принять во внимание факт, что все главные симптомы истерии могут быть вызваны путем простого внушения и путем внушения же устраняемы, то не имеется оснований сомневаться в психической природе этих симптомов. Но, с другой стороны, остаются еще некоторые проявления истерии, особенно со стороны растительных функций, которые трудно объяснить чисто психическим путем. Ввиду этого мы, руководясь физиолого-химическими исследованиями, склонны смотреть на истерию, как на болезнь обмена, при которой поражается вся вообще нервная система и в особенности ее психические отправления.
С другой стороны, состояние, описываемое под названием различного рода фобий, особенно Экиролем, Фальре, Вестфалем, Крафт-Эбингом, Баллем, Тамбурини, Маньяном, мной, Пит-ром и Режи и др., в последнее время некоторыми из французских автороз, особенно Жане, Реймоном и др., выделяется из обычных форм неврастении под названием психастении. Со своей стороны в русской литературе еще в 1885 г. мною была подробно описана та же форма болезни под названием психопатии или психоневрастении Ч
Надо при этом иметь в виду, что под формой собственно неврастении понимаются проявления раздражительной слабости и общего подавления нервной системы. При этом с психической стороны обнаруживаются элементарные симптомы, отражающие общее понижение нервной деятельности в виде быстрой психической утомляемости, внутреннего беспокойства и тоски, излиш-
1 См. В. Бехтерев, О психопатии и отношении ее к вопросу о вменении, сообщение в обществе врачей при Казанском университете, 1885.
406
ней склонности к развитию аффектов, неустойчивостью настроения и т. п. К психастении же, resp. психоневрастении, относятся те болезненные состояния, при которых вышеуказанные физические проявления отступают на второй план и в первую голову выдвигаются совершенно специальные психические расстройства в виде различного рода навязчивых состояний, так называемых фобий и одержимости, в виде поразительного безволия, беспричинной тоски и страха, систематических тиков и т. п. В этиологическом отношении также обе болезненные формы должны быть обособляемы друг от друга, так как первый невроз развивается обыкновенно у взрослых лиц от разнообразных случайных причин физического или нравственного характера, резко ослабляющих нервную систему. При этом имеется нередко и наследственное предрасположение, но оно большей частью незначительно и скрыто, тогда как психастения развивается обыкновенно у лиц, глубоко предрасположенных, приобревших дегенерацию от рождения или по наследству, причем черты ее обнаруживаются уже в сравнительно молодом возрасте.
Как бы то ни было, обе эти формы болезни и в особенности психоневрастения или психастения представляются не столько неврозами, сколько психозами и потому они в последнее время вполне по праву получают название психоневрозов х.
Что касается эпилепсии, то мы должны иметь в виду, что, наряду с большими приступами, в которых обнаруживается временная полная утрата нервно-психической деятельности, следовательно, симптом, несомненно, психического характера, и вместе с тем тонические клонические судороги, представляющие симптом нервного характера, имеются обыкновенно и приступы малой или так называемой психической эпилепсии, при которой все явления протекают в так называемой психической сфере. Если мы затем примем во внимание существование особых изменений характера у эпилептиков, частое развитие психической слабости и временное появление психических расстройств в форме так называемых эпилептических психозов, то станет понятным, что мы здесь имеем дело с таким неврозом, при котором чисто нервные симптомы нередко уступают по своей силе и по значению симптомам психического характера.
Кроме того, нельзя не принять во внимание, что настоящий психоз, как и психоневроз, сближаются между собой не только потому, что симптомокомплекстех и других представляет много общего между собой, но вместе с тем и потому, что общие психоневрозы даюг основание к развитию так называемых переходных или смешанных форм. И в самом деле, неврастения, психастения, меланхолия, ипохондрия и даже истерия в начале своего развития имеют вообще так много общего между со-
1 См. F# R а у га о η d, Nevroneset psychonevr Gses etc., Paris, 1907.
407
бой, что требуется нередко продолжительное наблюдение, чтобы окончательно остановиться на том или другом диагнозе.
Если мы затем углубимся в патогенез заболеваний, известных под названием психоневрозов или больших неврозов, то должны будем иметь в виду главным образом то или другое нарушение обмена, приводящее к изменению функций нервной системы вообще и мозговой коры в особенности. По крайней мере этот взгляд, основывающийся па исследованиях обмена веществ, в последнее время приобретает все более и более прав и по отношению к так называемой генуинной эпилепсии. Равным образом имеются ясные указания и на зависимость истерии и неврастении от нарушения в обмене веществ *.
С другой стороны, многие из психических расстройств, особенно меланхолия, мания, острая аменция и некоторые другие, также в основе своей имеют нарушение в обмене веществ, в пользу чего по крайней мере могут быть приведены многие данные.
Итак, не может быть сомнения, что психические болезни и общие нервные, а в частности, черепномозговые болезни в пато-лого-биологическом смысле представляют собой если не одно целое, то во всяком случае трудно обособляемые друг от друга области патологии, вследствие чего они нередко и изучаются в курсе медицинских наук в одной клинике, разделенной на два отделения, из которых одно служит для психических, а другое для нервных больных.
Переводя на практический язык все вышесказанное, можно утверждать без риска впасть в преувеличение, что нельзя претендовать на основательное изучение психических болезней без знакомства с нервными болезнями и, с другой стороны, нельзя быть хорошим невропатологом, не будучи знакомым и с психическими болезненными состояниями.
Тем не менее практическая важность обособления психических больных от больных всех других категорий и в том числе нервных приводит к необходимости выделения психических больных как от нервных, так и от иных соматических больных. В этом отношении мы должны сказать, что хотя и невозможно во многие случаях строго научное обособление психических болезненных состояний от нервных, но в практических видах все те случаи, где изменения психической сферы обнаруживаются в течение всего болезненного состояния и где они имеют преобладающее значение, можно признавать психическими болезнями, тогда как другие поражения нервной системы, в которых психические изменения играют меньшее значение по сравнению с нервными или представляются эпизодическими в течение болезненного состояния, могут и должны быть признаваемы нервными болезнями.!
'В. М. Бехтерев, О нарушении обмена веществ при неврасте пии, Обозрение психиатрии, 1898, стр. 527; Neurol. Zbl., 1898.
408
На том же основании и некоторые соматические заболевания, например, тифы и др., несмотря на частое обнаружение в течение болезни ясных симптомов со стороны нервно-психической сферы, не относятся к категории психических заболеваний.
Та тесная связь, которая существует между психическими и нервными болезнями, естественно, сближает психиатрию с остальной медициной, так как нервные болезни всегда стояли в тесной связи с другими медицинскими науками. Но не в меньшей, если еще не в большей мере психиатрию сближают с остальными медицинскими науками новейшие взгляды на происхождение многих психических болезней, как на результат самоотравления организма.
С этим вместе для огромного большинства психических заболеваний должно быть признано, что лежащее в основе их поражение мозговых функций не является первичным и самостоятельным, а обусловливается аутоинтокоикациями и действием токсических начал, поражающих весь вообще организм и в том числе нарушающих функции высших центров нервной системы и их сочетательных связей. Иначе говоря, дело не идет в этих случаях о первичном поражении собственно головного мозга, а о поражении всего организма; самые же психические болезни являются совокупностями функциональных расстройств или связанных с общими условиями жизнедеятельности организма и нарушением его отправлений в форме аутоинтоксикации того или другого рода, или связанных с влиянием общей инфекции и действием тех или других токсинов на мозг. В этих случаях изменения, происходящие в мозгу, могут быть' рассматриваемы лишь как производные основного поражения организма, имеющего свою этиологию, свое развитие, течение и даже исход в той мере, в какой на него не оказывают влияния те или другие терапевтические мероприятия.
Если такие органы, как центральная нервная система, при этом поражаются в большей степени, чем другие части организма, то это может обусловливаться самой инфекцией или аутоинтоксикацией и избирательным ее свойством подобно другим ядам, обладающим этой особенностью, далее, — чрезвычайной нежностью самой ткани мозга, особыми функциональными и иными условиями, ставящим мозг при данных условиях в положение locus minoris resistentiae, индивидуальными особенностями данного организма,приобретаемыми по наследству и условиями прошлой жизни и т. п.
При этом и те анатомические изменения в мозгу, которые открываются в такого рода случаях, лишь в известном числе являются непосредственными виновниками психических расстройств, чаще же они служат производными аутоинтоксикации, как и те психические расстройства, которые являются ее последствием. Дело идет в этом случае об изменении состава крови и развитии в ней химических продуктов и токсинов, которые, отравляя
409
нервную ткань, с одной стороны, вызывают в ней расстройство функциональной деятельности, а с другой, действуя в течение известного времени в виде более или менее длительных влияний, вызывают изменения мозговой ткани.
В этих случаях последние нам не могут служить объяснением болезненных симптомов, как изменения мозговой ткани при введении в организм внешних, так называемых интеллектуальных ядов не объясняет нам тех функциональных расстройств нервно-психической сфзры, которые имя обусловливаются. Дело в том, что интоксикации, как и аутоинтоксикации могут вызывать функциональные расстройства деятельности мозговых клеток, производя временные изменения в мозговом кровообращении и данамические изменения в питании клеток мозга без стойких структурных их изменений. Если впоследствии при хронических отравлениях в клетках и обнаруживаются уже более-стойкие структурные изменения, то они являются последствием продолжительного нарушения питания и отравления клеток и, следовательно, сами по себе как явления вторичные не могут быть принимаемы за основу функциональных расстройств психической сферы.
К этому следует добавить, что и изменения нервной ткани, производимые различными ядами, могут быть более или менее сходственными и даже одинаковыми, так как в сущности влияние ядэв не столько действует непосредственно на протоплазму клетки, сколько на условия ее питания, нарушение которых и приводит к более или менее одинаковым изменениям структуры клетки, несмотря на различие действия ядов.
Само собой разумеется, что упомянутые структурные изменения мозговых клеток, обусловленные аутоинтоксикациями, не безразличны для нервно-психических отправлений, но мы большей частью не можем с определенностью указать их значение в общей картине заболевания, пока дело не идет о невознаградимой атрофии нервных клеток и волокон, сопровождающейся развитием слабоумия.
Все вышеизложенное приводит нас к выводу, что корни душевных болезней, если исключить сравнительно редкие случаи психических расстройств, обусловленных травматическими повреждениями черепа, лежат не в головном мозгу, а в нарушении жизнедеятельности организма, обусловленном теми или иными внутренними дефектами или внесенными извне общими болезнетворными началами, причем от всех других болезненных состояний они отличаются лишь тем, что в результате нарушенной жизнедеятельности организма выдвигается на первый план по резкости внешних проявлений функциональное или органическое нарушение деятельности мозга, приводящее к психическим расстройствам. f|
Только что изложенное сближает психические болезни с общимл соматическими болезнями, так как здесь, как и в обык-
410
новенных болезненных процессах, дело идет также об общих аутоинтоксикациях или интоксикациях организма, влиянию которых подвергаются по преимуществу высшие центры головного мозга, заведующие психическими отправлениями, тогда как сопутствующие им соматические расстройства оказываются крайне слабо выраженными и мало заметными.
Из вышэизложенного очевидно, что следует совершенно отказаться от анатомического принципа в объяснении разнообразных психических расстройств, наблюдаемых при душевных болезнях. Как бы тонки ни были современные методы исследования нервной ткани мозга, они в большинстве случаев открывают нам в сущности лишь результаты действия на мозг аутоинтоксикаций и интоксикаций, которые, отравляя нервную ткань, собственно и лежат в основе психических расстройств, а не те анатомические изменения, которые являются результатом их воздействия на мэзг. Иначе говоря, основа душевных или психических болезней должна быть патолого-физиологическая, а не патологоанатомическая, причем последние если и играют роль в развитии психических расстройств, то лишь роль вторичного, а не первичного фактора. При этом, конечно, не умаляется и значение патологоанатомических изменений мозговой ткани, особенно в случаях, где как в органических психозах, они достигают крайне резкой степени своего развития и приводят к местным или разлитым разрушительным процессам, но тем не менее и в большинстве органических психозов патологоанатоми-ческие изменения мозговой ткани не составляют первичной основы патологического процесса, часто поражающего весь организм или по крайней мере коренящегося в поражении внечереп-ных органов.
Все вышесказанное должно существенным образом влиять, с одной стороны, на клиническое исследование и распознавание душевных болезней, с другой стороны, на их терапию.
Прежде всего необходимо отрешиться от того взгляда, что проявления психических болезней, как мы их наблюдаем в клинике, служат' выражением анатомических изменений мозга, и, следовательно, нет основания доискиваться и анатомической основы в разделении психозов, несмотря на сделанные в этом отношении попытки со стороны известных психиатров.
Сами по себе психические расстройства или симптомы в виде психического возбуждения, угнетения и безразличия, в виде иллюзий и галлюцинаций, в виде нарушений в закреплении и оживлении следов, в виде расстройств сочетательной деятельности, ненормальных влечений и т. п. еще не говорят нам о сущности данного заболевания, как и другие симптоматические состояния из области внутренних болезней, например, водянка, желтуха, кашель и т. п., не говорят нам о сущности того болезненного процесса, который лежит в их основе. Поэтому в симптоматическом отношении в известные периоды могут представ-
411
лять внешнее сходство совершенно различные душевные болезни, а потому дль правильной оценки психического расстройства ничуть не достаточно довольствоваться только наблюдением одних симптомов душевной болезни и выдвигать их как основу для диагностики душевных болезней, как это делается многими еще и по сие время.
Напротив того, только наблюдение всей совокупности симптомов, как психических, так и физических, та или другая смесь этих симптомов, их сочетание и течение, а равно и исследование всего организма с физической стороны и, между прочим, в отношении его наследственного и, в частности, семейного расположения, в отношении его возраста и телесного и умственного развития, в отношении присутствующих в организме отклонений или его индивидуальных особенностей, в отношении ранее перенесенных физических и психических болезней, а также бывших ранее излишеств того или иного рода или иных отягчающих моментов дают тот материал, при сопоставлении которого мы получаем возможность правильно оценивать данное психическое заболевание.
Если при этом остается невыясненным для нас действительный источник развития болезненного психического состояния, то вэ всяком случае, приняв во внимание все вышеуказанные данные, мы характеризуем всю совокупность условий и внешних особенностей данного болезненного состояния, причем можем быть уверены, что при совпадении этих условий в другом организме и при общем сходстве внешних проявлений психоза мы встретимся с одинаковым по существу психическим заболеванием, хотя бы его внешние проявления в зависимости от тех или других случайных моментов и представляли некоторые своеобразные особенности и отличия.
Что касается терапевтического вмешательства при психозах, то необходимо помнить, что все наши мероприятия, направленные к успокоению болезненных симптомов, к устранению бессонницы, отказа от пищи и т. п., суть лишь симптоматические средства, которые могут облегчать течение болезни, но не искоренять самую болезнь. Для последней цели необходимо все внимание направить на самую основу болезни и, где это возможно, попытаться ее устранить. Если же основа болезни остается скрытой или неустранимой, то необходимо позаботиться по крайней мере о лучшем питании и об улучшении обмена веществ находящимися в нашем распоряжении средствами, об исправном содержании желудочно-кишечного канала и т. п., чтобы дать больному организму самому лучше справиться с теми нарушениями в обмене веществ и с аутоинтоксикацией, которые лежат в основе данного психического заболевания.
Само собой разумеется, что правильное нозологическое выяснение душевных болезней должно соответственным образом влиять и на индивидуальную и общественную профилактику
412
подобного рода психических расстройств, так как последняя возможна только с распознаванием или по крайней мере предугадыванием истинной причины заболевания.
Равным образом и предсказание относительно дальнейшего течения и исхода душевной болезни стоит в прямой зависимости от правильного нозологического распознавания болезни, причем, однако, всякий раз должны быть приняты во внимание условия содержания и течения болезненного состояния. Дело в том, что каждая болезнь, предоставленная самой себе, должна иметь свое особое течение и исход. Те болезни, сущность которых находится во временном нарушении функций организма, состоящем в зависимости от каких-либо случайных внешних причин, обыкновенно заканчиваются по истечении известного времени полным выздоровлением и можно быть уверенным, что больному не грозит впредь новое заболевание, если, конечно, не повторятся те внешние причины, которые вызвали первоначальную болезнь; в других же случаях, где причины болезни коренятся в особых нарушениях обмена, связанных с периодически обостряющимся нарушением функционирования тех или других органов, мы имеем поразительную склонность болезни к возвратам, вследствие чего больному, освободившемуся от одного из приступов болезни, судьба грозит в будущем новыми и новыми болезненными приступами той или иной длительности.
В других случаях болезненное состояние, имеющее в своей основе неустранимое нарушение обмена и настоящую аутоинтоксикацию, коренящуюся не в случайных внешних причинах, а во внутренних условиях организма, роковым образом затягивается на более долгое время и заканчивается более или менее глубоким слабоумием. Наконец, в известном ряде случаев мы имеем дело с такой основой болезни, которая, будучи в корне неустранимой, постепенно все более и более подтачивает умственные и физические силы больного, причем болезнь, принимая прогрессирующее течение, заканчивается не только более или менее полным психическим распадом, но и летально.
Все эти различия в ходе и развитии душевных болезней стоят в прямой связи с истинной основой болезненного процесса и с условиями возникновения и дальнейшего развития, а не с внешними проявлениями отдельных симптомов, а потому ив профилактическом отношении должно играть большое значение распознавание истинной природы заболевания, а не внешних только проявлений болезни.
О МАНИАКАЛЬНО-МЕЛАНХОЛИЧЕСКОМ ПСИХОЗЕ
(К вопросу о соотношении между маниакальными и меланхолическими состояниями)х
Как известно, за последний период времени старое учение о мании и меланхолии, ведущее свое начало исстари, еще со времен Гиппократа, переживает свой кризис в том смысле, что Крепелин и его школа на основании клинических наблюдений стали отвергать существование прежде принимавшихся всеми авторами непериодических форм мании и меланхолии, признавая все вообще формы мании и меланхолии за периодические, образующие вместе с так называемым циркулярным психозом, альтернирующим и двойственным психозом (folie a double forme) одну общую клиническую форму маниакально-депрессивного психоза.
Правда, по отношению к меланхолии Крепелин первоначально был менее решительным, нежели по отношению к мании, так как до позднейшего времени в своей классификации удерживал еще так называемую инволюционную или климактерическую меланхолию как особый психоз непериодического характера, но с выходом работы Дрейфуса, нанесшей окончательный удар обособлению инволюционной меланхолии в виде самостоятельного психоза, и в этом отношении произошли изменения взглядов автора в том смысле, что все вообще формы меланхолии, как и все формы мании, должны быть признаваемы за периодические и относятся к маниакально-депрессивному психозу.
В сущности школа Крепелина, включая его многих последователей, в числе которых нельзя не упомянуть Шпехта, Виль-манса, Дрейфуса и др., все функциональные психозы сводит к двум большим клиническим формам: к dementia praecox с ее разновидностями и к маниакально-депрессивному психозу, к которому Вильманс относит, между прочим, и случаи циклотимии (Геккера)2. Но и между этими двумя формами оказалось так много
Ф 1 Доклад в научных собраниях клиники душевных и нервных бо-
лезней, 1908.
2 Как известно, Урштейн в своей монографии подвергает особенно резкой критике те основания, которые крепелиновской школой считаются достаточными для обособления dementia praecox от маниакально-депрессивного психоза.
4К
общего, что потребовалось немало усилий, чтобы провести и установить диференциально-диагностические признаки между той и другой1. При этом выяснилось, что не малое число случаев должно было войти в группу смешанных случаев маниакально депрессивного психоза, к которой, естественно, отошли многие из случаев dementia praecox с более благоприятным течением. Такому расширению понятия о маниакально-депрессивном психозе много содействовали работы Шпехта и Вильманса в 1907 г., а также исследования Дрейфуса, высказавшегося как мы видели, с решительностью против обособления так называемой инволюционной или климактерической формы меланхолии в особый психоз.
Шпехт в своей позднейшей работе2, которая вышла уже после известной работы Дрейфуса, настойчиво доказывает, вопреки взгляду Циена, Вестфаля и Кельпина, что инволюционная или климактерическая меланхолия относится также к маниако-депрессивному психозу. Автор приходит к этому выводу на основании анализа симптомов ажитированнои меланхолии, под видом каковой обыкновенно и протекает инволюционная меланхолия. Как известно, большинство авторов наблюдаемые при этой болезни симптомы возбуждения объясняли страхом, благодаря чему Вернике даже обозначал этот психоз именем Angst-melancholie (меланхолия страха). Но автор оспаривает это мнение, признавая, что ажитация здесь является результатом присоединения маниакального элемента к меланхолической картине, смесь же того и другого состояния именно и характерна для маниакально-депрессивного психоза, тогда как периодичность того и другого стоит уже на втором плане3.
Вышеуказанное наблюдение, ведущее нас к клиническому синтезу, имея на своей стороне многих горячих защитников4, до сих пор не остается без противников, к которым следует отнести Жолли, Циена, отчасти Вестфаля, Кельпина и целый ряд других авторов, высказывающихся частью против самого учения о маниакально-депрессивном психозе, частью только
1 По Шпехту, даже и paranoia chronica — этот типический психоз — должен быть отнесен к маниако-депрессивному психозу, так как в нем будто бы можно найти все симптомы мании, как повышенное самочувствие, суетливое беспокойство, стремление писать и т. п., и так как, с другой стороны, существуют переходные формы между маниако-де-прессивным психозом. Нечего и говорить, что взгляды эти встречают известную оппозицию в психиатрической литературе, о чем нет надобности здесь распространяться.
2 S р е с h t, Zschr. f. Nerv. u. Psych., 1908, № 12.
3 Заслуживает внимания, что, по указаниям Шпехта, Гален еще в 1868 г. признавал родство меланхолии с манией и признавал маниакально-меланхолические смешанные состояния (Mischzustande), но его взгляды не получили признания со стороны современников и были вскоре забыты.
4 Между последними должно отметить и Дени и Камю — авторов недавно появившейся монографии о маниакально-депрессивном психозе.
415
против его расширения, допускаемого в последнее вр,емя школой Крепелина.
В числе последних исследований, относящихся к занимающему нас вопросу, нельзя не отметить работы Урштейна1, который восстает главным образом против смешанных состояний. Он находит, что виной всего этого учения о смешанных состояниях (Mischzustande) является переоценка маниакально-депрессивных /"симптомов и умаление кататонического сим-птомокомплекса..»
Автор держится того взгляда, что маниакально-депрессивные симптомы могут наблюдаться при всевозможных психозах и тем не менее они ничуть не могут исключать этих психозов, как и dementia praecox. Так, у целого ряда больных, дошедших до состояния кататонического слабоумия, автор описывает явления, считающиеся характерными для маниакально-депрессивного психоза, причем состояние душевной пустот ы(интрапси-хического торможения) автор даже признает характерным для dementia praecox. С другой стороны, он находит, что ни периодичность, ни богатая смена возбуждения и угнетения не дают основания признавать существование особого маниакально-депрессивного психоза.
Точно так же автор отвергает и взгляд Дрейфуса об исключении старческой меланхолии из числа особых психозов, кроме случаев, относящихся к поздним формам dementia praecox.
Мы не будем здесь входить в подробности других многочисленных литературных разноречий по вопросу о маниако-депрес-сивном психозе, которые и до сих пор не могут считаться окончательно устраненными.
Заметим лишь, что обширная литература поэтому вопросу, полная и до сих пор еще страстных споров, без сомнения, указывает, что развитие учения о маниакально-депрессивном психозе еще не завершило своего естественного цикла, а потому нам кажется нелишним еще раз остановиться на некоторых спорных вопросах этого учения.
Первый вопрос, связанный с учением о маниакально-депрессивном психозе, который возбуждал много споров, это — вопрос об отсутствии непериодических форм мании и меланхолии, до сих пор не может считаться окончательно решенным особенно для меланхолии.
Бесспорно, что периодические формы меланхолии и мании встречаются значительно чаще, нежели непериодические, но все же последние, повидимому, не могут быть исключаемы совершенно.
Дело в том, что в некоторых случаях и мания, и меланхолия, развиваясь впервые в инволюционном периоде или даже в мо-
1 U r s t e i η, Die Dementia praecox und ihre Stellung zum manisch-dcpressiven Irresein, 1909.
416
лодом возрасте, не обнаруживают никакой склонности к повторным возвратам и случается, что больной, заболев однажды меланхолической формой, доживает до глубокой старости, не обнаружив никаких признаков возобновления психоза в последующей жизни.
В других случаях впервые психоз в форме меланхолии или мании обнаруживается в молодом возрасте и затем не возобновляется вовсе в течение всей последующей жизни или же если и возобновляется, то иногда лишь в другом критическом возрасте, например, в инволюционном периоде, во время климактерия, следовательно, спустя 20—30 лет после первого заболевания, причем психоз остается иногда неизлеченным, а если протекает благоприятно, то после этого может уже не возобновиться совершенно.
В таких случаях может ли быть речь о периодичности психоза в настоящем смысле слова и неуместно ли было бы говорить лишь об известном предрасположении к развитию психоза рецидивированию его, а не о периодическом психозе.
Очевидно, что, не прибегая к натяжкам, трудно говорить в этих случаях о периодическом психозе по самому существу, так как в противном случае понятие периодичности утрачивает всякое ограничение.
Действительно, были даже попытки утверждать для тех случаев, когда психоз развился однажды и не возобновлялся в течение всей дальнейшей жизни человека, что в данном случае периодичность его была пресечена смертью, что, если бы не наступила смерть, психоз наверно бы возобновился. Но такие утверждения, во-первых, без греха против логики можно применить и к другим излечимым психозам, во-вторых, эти утверждения совершенно не считаются с возможностью настолько прочного улучшения в деятельности нервно-психических функций организма после однажды перенесенного психоза, что для возобновления того же психоза уже не имеется благоприятной почвы.
Мы не будем здесь останавливаться на клинических проявлениях периодических психозов, которые, как известно, отличаются нередко некоторыми особенностями, дающими возможность отличать эти психозы от непериодических психозов.
Сюда относятся такие особенности клинического течения периодических психозов, как возникновение отдельных приступов болезни без особых внешних поводов, сходство отдельных приступов в развитии и течении, повторность приступов через более или менее правильные промежутки времени, быстрое начало и быстрое обрывание приступов, относительная краткость отдельных приступов, существование коротких просветлений в течение приступов и неполнота нервно-психического здоровья в промежутках между приступами периодических форм и, нако-
27 В. М. Бехтерев
417
нец, существование абортивных кратковременных приступов, психоза в течение так называемых светлых промежутков и т. п.
Дело в том, что эти признаки, как упомянуто, дающие возможность в известных случаях клинически различать периодические психические расстройства от непериодических, сами по себе, по утверждению некоторых авторов, не могут считаться настолько постоянными, чтобы могли руководить нас везде и всюду в распознавании периодических психозов.
Таким образом, и этот признак, который мог бы нас руководить в решении вопроса о периодичности мании и меланхолии, отпадает. Но если с новым учением совершенно стираются те основные признаки, по которым ранее обособлялись периодические психозы от непериодических (повторность и клинические особенности), то не подлежит сомнению, что новое учение не внесло в науку и убедительных данных в пользу всегдашней периодичности мании и особенно меланхолии.
Как известно, и ряд статистических исследований, предпринятых различными авторами, между которыми мы упомянем работы Майзера, Шотта, Липшица, Келышна, Сэза, Суханова и Ганнушкина, Ламбропци, Гинриксена, Кауппа, Пейксото, Пераццоло, Гуччи, Рама, Уокера и др., не решает этого вопроса в положительном смысле.
Напротив того, в вышеуказанных статистических работах отмечается с постоянством тот или другой, во всяком случае не малый процент однократных заболеваний манией и меланхолией.
Отсюда, казалось бы, можно было сделать вывод, что не всегда маниакальные и меланхолические формы протекают периодически, что имеются и непериодические формы тех же психозов. При этом, однако, несомненно, что как меланхолия, так и мания в значительном числе случаев развивается у лиц, носящих в своем организме известное предрасположение к повторному заболеванию.
Признав этот факт, мы не имеем в то же время никаких оснований делать различия по существу между периодическими и непериодическими формами меланхолии и мании.
Как известно, многие авторы в своих классификациях душевных болезней периодические психозы выделяли от обыкновенных в особую группу, основываясь, очевидно, на существенных различиях между теми и другими в смысле этиологии, развития, течения и предсказания.
Эта точка зрения мне казалась всегда ошибочной, в силу чего в классификации душевных болезней, вышедшей еще в 1887 г., периодические формы мании и меланхолии были отнесены мною к той же группе, в которой помещены и обыкновенные психозы г.
1 С тех пор, как вышла вышеуказанная моя брошюра о классификации душевных болезней, психиатрия сделала такие успехи, что, само
418
Это объединение периодических и непериодических психозов в одной группе имеет в виду то предположение, что в основе и тех, и других психозов лежит одно и то же болезнетворное начало с тем лишь различием, что при периодическом психозе" оно присутствует непрерывно в организме, причем его действие -затихает лишь временно и затем вспыхивает вновь, тогда как-при непериодическом психозе основное болезненное начало-обнаружило при известных условиях свое действие на организм, но затем происшедшее расстройство компенсировалось-в такой мере, что оно уже не в состоянии проявлять своего-действия по крайней мере до тех пор, пока не наступят какие-либо сильные внешние влияния, которые могут вновь вывести организм из равновесия.
Ясно, что если в периодических и непериодических психозах мы вынуждены признавать однородное болезнетворное начало-с тем лишь различием, что в одних случаях оно не исчезает бесследно из организма, а вновь заявляет о себе в известные периоды времени, тогда как в других случаях оно возникает в организме лишь временно при известных условиях и затем компенсируется более или менее окончательно, то не может быть различия между теми и другими психозами по существу, а дело идет лишь о различии в смысле более стойкого и менее стойкого расположения к болезненным проявлениям.
Таким образом, хотя и не исключается нами существование непериодических форм меланхолии и мании, но мы высказываемся в смысле объединения периодических и непериодических форм мании и меланхолии, не придавая вопросу о периодичности того значения, которое ему часто приписывается авторами.
С другой стороны, как известно, благодаря учению Крепе-лина ныне устанавливается объединение меланхолических и маниакальных форм между собой. Уже и прежде, как известно, меланхолия и мания вмещались обыкновенно в одну группу острых психозов или психоневрозов по Крафт-Эбингу, но Кре-пелин объединил прежние состояния меланхолии и мании, рассматривавшиеся, как самостоятельные формы, в одно целое вместе с циркулярными психозами под общим названием маниакально-депрессивного психоза.
собой разумеется, эта классификация не может уже удовлетворять и моим взглядам. В частности, I и II группы психозов, на мой взгляд, могли'бы быть соединены в одну группу под названием «простые психозы», delirium ncutum как инфекционную болезнь, патологическая анатомия которой ныне изучена много лучше прежнего, должно отнести к органическим психозам. Кататония и гебефрения должны быть объединены под общим; наименованием dementia praecox и помещены в психозах вырождения» круговое помешательство должно быть объединено с манией и меланхолией под общим названием маниакально-меланхолического психоза и должно помещаться в первой группе вместе со своими исходными состояниями. О других изменениях, которые мы считали бы необходимым ввести в нашу классификацию, мы здесь распространяться не будем, имея, в виду сказать об этом в другом месте.
27*
419
Основанием для этого, кроме отрицания непериодических форм мании и меланхолии, послужило существование переходных форм между манией и меланхолией. Хотя на первый взгляд и кажется, что меланхолия и мания представляют противоположности друг другу, но в действительности между теми и другими имеются переходные состояния, известные под названием смешанных форм.
К этим смешанным формам относится, между прочим, известная уже издавна «гневная мания», где, наряду с ускорением . ассоциаций и двигательным возбуждением, имеется не столько веселое расположение, сколько раздражительность, гневливость и недовольство окружающим. К таким же смешанным формам относится так называемая депрессивная мания, где имеется двигательное возбуждение и где одновременно имеется отрицательное колебание чувственного тона, причем поражает бедность и однообразие ассоциаций. Далее, сюда же относится так называемая «непродуктивная мания», т. е. мания, не сопровождающаяся быстрой сменой ассоциаций, где, по Кре-пелину, дело идет даже о замедленном восприятии и образовании сложных концепций. В этих формах можно видеть как бы сближение мании с меланхолией. Но имеются и другие смешанные формы, которые сближают меланхолию с манией. К ним относится, по Крепелину, «маниакальный ступор», т. е. состояние, когда при оцепенелости у больных обнаруживается улыбка или ирония. Сюда же относится «депрессивное состояние с ускорением в течении идей», когда у больных при дурном настроении происходит такой наплыв мыслей, что больные по их заявлению не могут ими управлять, причем здесь наплыв мыслей не может лишь выражаться внешним образом, вследствие задержки в речевом аппарате. Кроме этих форм, Крепелин рассматривает еще смешанную форму,, которую он называет «маниакальной задержкой» и при которой, наряду с хорошим настроением и облегченным ходом ассоциаций, имеется психомоторная задержка. Дело идет здесь, следовательно, о маниакальном состоянии без двигательного возбуждения.
Эти смешанные состояния признаются как промежуточные или переходные формы между меланхолией и манией, благодаря "чему та и другая как бы связываются между собой множеством переходных форм. К этим промежуточным формам можно было ■бы присоединить еще состояние, известное под названием так называемой «активной меланхолии», где имеется резкое угнетенное состояние и одновременно двигательное возбуждение и, ■с другой стороны, всем известный raptus melancholicus, являющийся примером аффекта, при котором совмещается отрицательный тон с резким двигательным возбуждением. Но возникает вопрос, имеют ли эти смешанные состояния на самом деле то значение, которое им приписывает школа Крепелина.
Выше мы видели, что Шпехт в вопросе о маниакально-депрес-
420
сивном психозе существование смешанных состояний выдвигает даже на первый план, периодичность же ставит на втором плане и тем самым считает возможным отпарировать возражения, опирающиеся на существование непериодических форм меланхолии и мании.
Таким образом, очевидно, что эти смешанные состояния играют особо видную роль в объединении мании и меланхолии в одну клиническую форму под названием маниакально-депрессивного психоза.
Однако что могут доказывать переходные формы в таком частном вопросе, как вопрос о родстве между меланхолией и манией.
Дело в том, что переходные, resp. смешанные, формы имеются везде и всюду, так как природа вообще не делает скачков. Какой бы психоз мы вообще не взяли, мы всегда найдем множество переходных форм между ним и другими психозами.
Не подлежит сомнению, что имеются переходные формы даже между меланхолическим психозом и параноей, что уже' и привело некоторых последователей нового учения к отрицанию самостоятельности столь типического психоза, как хроническая параноя. Нетрудно затем указать и переходные формы между тем же маниакально-депрессивным психозом и dementia praecox, чему свидетельством служат те затруднения в диагнозе, с которыми мы иногда в этих случаях встречаемся. Равным образом нетрудно указать на переходные формы между amentia Meynertia и галлюцинаторной формой dementia praecox, между сифилитическим психозом и dementia paralytica, между эпилепсией и истерией и пр. Да и может ли быть иначе, если дело идет в этих случаях собственно о реакциях больного организма на болезнетворные начала, а не о различии этих самых начал. Таким образом, развивая далее учение о переходных или смешанных формах, мы могли бы установить известное родство' даже между далеко отстоящими друг от друга болезненными формами.
По изложенным основаниям мы не считаем в какой-либо мере особенно убедительным доказательством существование смешанных или переходных маниакально-меланхолических состояний в пользу признания маниакально-депрессивного психоза. Гораздо большее значение, по нашему мнению, имеют другие клинические данные и факты, которые не оставляют сомнения в близком соотношении маниакального и меланхолического состояния.
Одно из существенных данных, говорящих в пользу вышеуказанного соотношения, мы видим в клинически часто констатируемом переходе или превращении состояний угнетения в состояние возбуждения и, наоборот, даже при непериодическом течении болезненного состояния с характером мании или меланхолии. Так, уже давно известно, что маниакальный психоз
42 г
начинается почти обычно стадией угнетения, которое может быть более или менее резко выраженным и которое может быть различной длительности.
В отдельных случаях стадия угнетения может затягиваться на более или менее продолжительное время, а иногда, как я мог наблюдать в отдельных случаях, период угнетения даже затягивался на несколько месяцев. В этом случае собственно психоз протекал в форме двух фаз — меланхолической и#мани-акальной, причем и та, и другая фаза продолжались в течение нескольких месяцев, после чего психоз проходил окончательно, не обнаруживая тенденции к возобновлению приступов, как это наблюдается в случаях folie a double forme. Дело шло, таким образом, о двойственном психозе непериодического характера, в котором фаза меланхолическая сменялась маниакальной фазой.
В других случаях, как известно, вслед за маниакальной фазой психоза следует фаза угнетения или утомления, как называли ее прежде. В этом случае вслед за прекращением маниакального состояния больной не сразу переходил в нормальное состояние, а перед выздоровлением обнаруживался более или менее длительный период угнетения, пока больной окончательно не переходил в выздоровление. И здесь должно отметить случаи, в которых вслед за маниакальными приступами следовал продолжительный приступ меланхолического угнетения, за которым уже наступало окончательное выздоровление, хотя опять-таки дело шло о психозе, не сопровождающемся повторением приступов, состоящих из двух фаз, как это мы имеем в случаях folie a double forme. С другой стороны, в отдельных случаях и после тяжелых меланхолических состояний перед окончательным выздоровлением больных наблюдается иногда очень ясно выраженная стадия маниакального оживления, степень и продолжительность которой представляются в отдельных случаях не одинаковыми.
Далее, в случаях повторного развития приступов мании, т. е. при несомненно периодической мании мы встречаемся иногда со случаями, где в промежутках между приступами мании может оказаться приступ меланхолического состояния и, наоборот, в случаях повторного развития приступов меланхолии, resp. при несомненно периодической меланхолии между приступами с меланхолическим угнетением в тот или другой период времени, может оказаться приступ мании. Наконец, чередующийся психоз (folie alterne typique), двойственный периодический психоз (folie a double forme) и круговой психоз (folie cir-cularie) снова представляют собой периодическую смену маниакального и меланхолического состояния, но в уже более явной и для всех очевидной форме.
И в этих формах можно почерпнуть многое достойное внимания для выяснения близкого родства между меланхолическими и маниакальными состояниями. Так, останавливаясь прежде
4,22
всего на чередующемся (folie alterne) и двойственном психозе (folie a double forme), необходимо иметь в виду, что далеко не всегда дело обстоит с приступами столь правильно, как это обыкновенно принято думать. В случаях folie alterne typique возможны случаи, когда правильная смена приступов возбуждения и приступов угнетения со' свободными между ними промежутками нарушается благодаря тому, что иногда следуют один за другим два приступа возбуждения или два приступа угнетения, разделяемые свободными промежутками, после чего снова устанавливается правильное чередование приступов угнетения и возбуждения.
Далее, при двойственном психозе (folie a double forme) мы встречаем прежде всего чередование приступов в таком виде, что первоначально наступает маниакальное состояние, за которым следует период угнетения и затем свободный промежуток времени, в других же случаях дело происходит как раз наоборот, иначе говоря, приступ начинается меланхолическим состоянием и сменяется затем маниакальным, после чего уже наступает свободный промежуток времени. Нам всем известно, что в развитии приступов этого психоза наблюдаются те или другие неправильности. Так возможно, что вслед за приступом возбуждения и угнетения может последовать непосредственно вновь приступ возбуждения и затем уже наступает светлый промежуток или, наоборот, дело начинается иногда периодом угнетения, за которым следует период возбуждения, а за последним вновь непосредственно следует угнетение, сменяющееся светлым промежутком. Иногда же в промежутках между двойными приступами следует одиночный сильно затянувшийся приступ возбуждения или же одиночный приступ угнетения.
Нечего говорить, что ни длительность отдельных периодов возбуждения и угнетения, ни длительность светлых промежутков между приступами также большей частью не отличаются большой правильностью.
Заслуживает также внимания Факт, что как в двойном психозе, так и в циркулярном пс^'озе обыкновенно чем резче бывает маниакальный приступ, тем глубже и психическое угнетение в последующем периоде и наоборот.
С другой стороны, известно также, что чем более значительна степень возбуждения и угнетения в отдельных приступах, тем они бывают в общем короче и, наоборот, чем менее они бывают выражены, тем они бывают более длительными.
Наконец, не лишена известного значения и быстрота смены одного состояния другим, которая происходит иногда в течение не более нескольких часов.
Все это такие клинические факты, которые гораздо более, нежели так называемые смешанные состояния, с нашей точки зрения, далеко не заслуживающие того внимания, которое им уделяют позднейшие авторы, доказывают как клиниче-
423
ское, так и генетическое родство между собой и последнее даже больше, нежели первое.
Выше уже была речь о том, что в вопросе об этом родстве не имеет существенного значения и признание или непризнание непериодических форм мании и меланхолии.
Ввиду вышеприведенных фактов не остается сомнения в том, что как для непериодических и периодических форм мании, так и для непериодических и периодических форм меланхолии должно быть принято одно болезнетворное начало с тем лишь различием, что в случаях непериодического течения психоза условия для проявления действия болезнетворного начала развиваются под влиянием того или иного случайного толчка, тогда как в случаях периодической мании и меланхолии в организме имеются условия для периодического возобновления действия того же болезнетворного начала, с другой стороны, и для сменяющихся состояний мании и меланхолии, как круговой психоз и его разновидности, а следовательно, и вообще для мании и меланхолии должно быть признано одно болезнетворное начало. Иначе, т. е. становясь на точку зрения двух болезнетворных начал, одного для мании и другого для меланхолии, нельзя понять, вследствие каких условий в организме происходит своеобразная игра двух болезнетворных начал, из которых одно в известных случаях не только сменяет другое, но и сообразуется с первым по силе действия и даже по продолжительности действия и притом сменяет часто непосредственно, так что одно болезненное состояние переходит в другое иногда в течение нескольких часов, а иногда и того менее. Напротив того, приняв и для маниакальных, и для меланхолических приступов одно и то же болезнетворное начало в организме, смену одного состояния другим, мы можем поставить в зависимость от степени действия болезнетворного начала.
Мы знаем, что действие всякого яда на нервную систему сводится к ее возбуждению и затем к последовательному развитию угнетения и паралича, но период возбуждения может оказаться слишком коротким и потому кажется иногда, как будто яд сразу приводит к развитию паралича, тогда как и в этих случаях дело сеодится к тому, что яд первоначально действует возбуждающим образом, а затем угнетающим. При этом современная наука не оставляет сомнения в том, что и возбуждение, и угнетение зависят в сущности от дозы ядовитого вещества: при малой дозе и сильный яд, обычно приводящий к угнетению и параличу, может приводить к возбуждению и даже действие его будет ограничено одним возбуждающим влиянием; наоборот, зараз введенная большая доза ядовитого вещества может привести почти сразу к развитию паралича. Приняв эти факты ео внимание, мы будем иметь полную возможность вполне удовлетворительно объяснить соотношение между меланхолией и манией, допустив гипотезу самоотравле-
4.'М
ния организма при этих болезненных состояниях, в пользу чего, мне кажется, в настоящее время имеется более чем достаточное число фактов.
В этом отношении уже кое-что дают нам физиолого-химиче-ские исследования. Так, известно, что при маниако-депрессивных состояниях находят как обычное явление увеличенное количество индоксила в моче. Люис К. Брюс х признает, что имеется связь между избытком индоксила в моче и состоянием депрессии и что индоксил является главным этиологическим моментом душевных заболеваний.
Надо, впрочем, заметить, что при исследованиях самого же-Брюса в 4 случаях меланхолии не было найдено увеличения индоксила в моче. Также и Кэвиет2 особенно часто находил увеличение индоксила в маниако-депрессивных состояниях, где, по наблюдениям автора, имеется настоящий параллелизм между развитием приступа и колебаниями индоксила. Следует иметь в виду, что индоксил открывается и в стадии угнетения, и в стадии возбуждения с тем лишь различием, что в депрессивной стадии его больше, чем в стадии возбуждения.
Само собой разумеется, что известная часть индоксила при этом может объясняться застоями в кишечнике, обусловливаясь происходящим в нем гниением вместе с развитием микроорганизмов, так как этот способ происхождения индоксила уже доказан и экспериментальным путем.
Так именно и смотрят на происхождение индоксила некоторые из авторов. По Пардо, увеличение индоксила доказывает только наличность бактерий, что зависит от кишечных застоев и функциональной недостаточности кишечника. Его собственные наблюдения, произведенные на 118 душевнобольных, показывают, что увеличенное количество индоксила в моче может быть поставлено в связь главным образом с известными стадиями в развитии душевного расстройства. При этом уменьшение индоксила совпадало с улучшением душевного расстройства; с другой стороны, при устранении антисептическими приемами индоксилурии происходило и улучшение психического расстройства; в случае же, если антисептика не оказывала влияния, не изменялось и психическое расстройство. Но такое объяснение увеличенного количества индоксила может иметь значение главным образом для депрессивной стадии психоза, в которой обычны кишечные застои; но ничего такого обычно не наблюдается в маниакальной стадии, а между тем при этом все же открывается увеличенное количество индоксила.
Заслуживает также внимания факт, что и своевременное очищение кишечника в депрессивной стадии не устраняет совершенно индоксила.
1 Bruce Lewis С, The clinical significance of indoxyl in the urine, The Journ. of ment. science, 1906, v. LII, № 218.
2 С a v i a t, Ar.icr. journ. of insanity, 1907.
425-
Это наводит на предположение, что увеличение индоксила 'при маниакально депрессивном психозе может иметь и другое происхождение, представляясь результатом нарушенного обмена веществ 1.
Известно, что белок в организме под влиянием протеоли-тических энзим расщепляется на ряд соединений, в числе которых, между прочим, имеется и триптофан (протеинохромоген), образующийся и при гниении белковых тел. При восстановлении триптофана в свою очередь распадается на а-амидо-про-теиновую кислоту и индол; последний же, окисляясь в организме, дает индоксил, который, будучи неустойчивым телом, дает при .дальнейшем окислении два изомерных вещества — индиготин и индирубин, а при большей степени окисления — изатин.
В организме индоксил связывается частью серной кислотой, частью гликуроновой и служит к образованию индоксил-серной кислоты и индоксил-гликуроновой2, но последняя, повидимому, образуется лишь вместе с чрезмерным образованием в организме индоксила, когда для его связывания серной кислоты оказывается уже недостаточно. Обе кислоты и появляются в моче, в которой путем прибавления крепкой кислоты они выделяют индоксил, превращающийся затем путем окисления (с помощью хлора, полуторахлористого железа и пр.) в гелиндиготин, а последний, благодаря своей неустойчивости, легко переходит в синий индиготин или красный индирубин, при избытке же окисления получается бесцветный изатин (Майяр).
Из вышеизложенного ясно, что индоксил может быть не только результатом гниения в кишечнике или гнойных процессов в организме, но что он может быть и результатом неправильного обмена веществ.
В пользу этого говорят, между прочим, и наблюдения Клеч-ковского и Блюменталя, которые доказывают увеличение индо-ксилурии при раздражении спинного мозга и его нервов и при сахарном уколе в продолговатый мозг. Поэтому естественно, что у душевнобольных индоксилурия может быть тканевого происхождения.
В пользу того же говорят также и исследования М. Кауфмана3 над эпилептиками, который наблюдал увеличение индоксила в моче перед припадком, после же припадка количество его уменьшалось. Тот же автор в двух случаях прогрессивного паралича констатировал, что, несмотря на голодание и тщательную промывку кишечника, количество индоксила в моче оставалось без изменения.
Можно предположить, таким образом, что при нарушении ■обмена веществ в самих тканях образуется триптофан, который и
1 Словцо в, Русский врач, 1907, № 7.
2 Schmiedeberg, Arch. f. exper. Path. u. Pharm., 14.
3 M. К a u f m a π η, Beitrage z. Pathologie d. Stoffwechsels bei Psv--chosen, 1908, Bd. 1.
42G
окисляется затем в индоксил. Есть возможность и клиническими методами выяснить происхождение индикана.
Так, при индоксилурии кишечного характера обнаруживается зависимость его от количества принимаемой пищи, увеличение при приеме щелочей, уменьшение при молочнокислой диэте и при .назначениях антисептических и mag. bismuthi.
При индоксилурии гнойного и тканевого происхождения ни приемы пищи, ни введение mag. bismuthi не изменяют его содержания в моче; но зато при тканевой индоксилурии поглощение щелочей уменьшает выделение индоксила, тогда как при гное индоксилурия не уменьшается вместе с введением щелочей.
Таким образом, и по отношению к вопросу о происхождении индоксила при маниакально-депрессивном психозе могут быть сделаны соответствующие разъяснения.
Действительно Зейге1 наблюдал увеличение индоксила при ■маниакально-депрессивном психозе, особенно значительное в периоды депрессии, несмотря на тщательное опорожнение кишечника с помощью карлсбадских .препаратов, вследствие чего ин-диканурию в этом случае автор относит к влиянию центральной нервной системы.
Позднее тот же автор2 на двух меланхоликах и трех больных циркулярным психозом также наблюдал большое количество индикана; при антисептическом содержании кишечника количество индикана достигало 0,5, с выздоровлением же оно падало резко..
В этом отношении заслуживает также внимания недавно произведенные у нас исследования д-ра Оморокова над определением количества индоксила у душевнобольных с целью выяснить, с одной стороны, связь индоксилурии с психическим состоянием, с другой — происхождение индоксилурии при психозах. Определение производилось по методу Валовского3, в котором показателем количеств индоксила являются цифры хлора, потребленного при окислении всего мочевого индоксила, причем случаи с потратой хлора большей 0,16 должны считаться повышенным содержанием индоксила. Другим методом для определения количеств индоксила служил метод Ванг-Баума, который исключительно затем и применялся. Исследования произведены над 21 больным, из которых 7 с маниако-депрессивным психозом, один случай старческой меланхолии, 2 с amentia, 4 с dementia ргаесох и 7 с прогрессивным параличом.
При этом оказалось, что в случаях dementia ргаесох увеличения индоксила не имелось. Также у двух аментиков не было
1 М. S e i g e, Periodische Indicanurie bei zirkularer Psychose, Mo-natcshr. f. Psych, u. Neurol., Bd. XXIV, Hft. 2.
2 Bericht v. d. Jahresamml. d. Vereins f. Psych., Monatschr. f. Psych., Bd. XXV, Hft. 6.
3 Соболев, Диссертация, СПБ, 1907; Тер-Григорианц, СПБ, }907.
427
особенно характерных изменений в отношении выделения индоксила. Напротив того, в 7 случаях прогрессивного паралича было постоянное увеличение индоксила, причем молочнокислая диэта, каломель и висмут понижали здесь количество индоксила до минимума, что доказывает кишечное его происхождение. В других 8 случаях прогрессивного паралича, исследованных время от времени, повышения индоксила не обнаружено.
Напротив того, маниакально-депрессивные состояния дали особенно интересные данные. Во всех 8 случаях обнаружено большое количество индоксила в моче, при этом в некоторых случаях этой формы повышенное количество индоксила, несомненно, тканевого происхождения, так как не уменьшалось ни от молочнокислой пиэты, ни от кломеля и висмута.
Таким образом, не относя происхождение маниакально-меланхолического психоза на счет увеличенного образования индоксила, необходимо признать, что у этих больных дело идет об аутоинтоксикационных процессах с таким изменением обмена, при котором белок распадается на ряд межуточных веществ, которые и способствуют повышению индоксила. Должно иметь в виду, что повышенные количества индоксила в организме сопутствуют ослаблению процессов окисления в организме, как показывают в случаях автора коэфициенты Робина.
Таким образом, эти данные не оставляют сомнения в том, что в организме больных с маниакально-меланхолическим психозом имеются нарушения обмена, зависящие от аутоинтоксикации, не объясняемой одной задержкой отправления кишечника, причем эта аутоинтоксикация, очевидно, сильнее выражена в депрессивной стадии, нежели в маниакальной.
В согласии с вышеизложенным стоят и исследования над газообменом, которые также доказывают понижение окислительных процессов при маниакально-меланхолическом психозе. Исследования, которые в этом отношении производились у нас д-ром Омороковым с аппаратом Гепперта и Цунца, показали, что задержка газообмена обнаруживается как в депрессивной стадии, так и в стадии возбуждения, причем понижение окислительных процессов происходит значительно резче, нежели в стадии возбуждения х.
Таким образом, становясь на точку зрения самоотравления организма как основы маниакально-депрессивного психозами руководясь вышеприведенными данными, необходимо прийти к выводу, что сущность самоотравления при этом психозе, с одной стороны, не обусловливается состоянием кишечника и, с другой стороны, представляется одной и той же как в меланхолической, или депрессивной, так и в маниакальной, или экзальтационной, стадии.
1 Омороков, Диссертация, СПБ, 1909.
42S
Но в меланхолической стадии самоотравление достигает большей степени, нежели в маниакальной. Таким образом, рассматривая две стадии маниакально-меланхолического психоза, маниакальную и меланхолическую, мы признаем для них одинаковый по сущности патогенез, состоящий в специфическом самоотравлении организма, но в маниакальной стадии, которая состоит в возбуждении функций мозговых центров, степень отравления меньше, в меланхолической же стадии, состоящей в угнетении функций нервной системы, степень отравления оказывается более значительной.
Само собой разумеется, что гипотеза самоотравления ничуть не исключает вазомоторной теории по отношению к этим психическим состояниям, которую проповедывал еще Мейнерт и которая в последнее время также развивается некоторыми из авторов, например, Таблитцером. Напротив того, следует допустить, что вступление в круг кровообращения вредоносного или ядовитого вещества, действующего в больном организме на сосу-додвигательную систему, и может обусловить 'в одних случаях спазм артериальных сосудов тела, в том числе и мозговых, в других случаях, наоборот, расширение артериальных сосудов.
Такая вазомоторная теория, которую принимаем и мы для объяснения патологических состояний угнетения и возбуждения, по нашему мнению, может вполне удовлетворительно объяснить как тесное клиническое соотношение маланхолии и мании, так и быструю смену в известных случаях одного состояния другим, а также известное соотношение в степени и глубине маниакальных и меланхолических приступов при двойственном (folie a double forme) и циркулярном психозах.
Если, таким образом, представляется возможным установить патогенетическое родство между манией и меланхолией, объясняющее нам и тесное клиническое соотношение между обоими состояниями, то отсюда очевидно, что оба эти состояния, несмотря на кажущееся внешнее различие и до известной степени даже ■противоположность явлений, должны быть обобщаемы под одним общим наименованием маниакально-меланхолического психоза.
С этим вместе различаемые ранее формы меланхолии простой и периодической, мании простой и периодической, кругового психоза, двойственного и чередующегося психоза могут быть рассматриваемы как разновидности одной общей клинической ■формы — маниакально-меланхолического психоза.
С вышеизложенной точки зрения становится совершенно излишним спор о том, могут ли встречаться мания или меланхолия не периодически. Отчего, спрашивается, не могут, если в одних случаях условия для самоотравления организма развились лишь под влиянием каких-либо случайных причин, затем более уже не повторяющихся, тогда как в других случаях в самом организме лежат условия для возобновления вышеуказанных условий,
429
приводящих к развитию самоотравления, причем в зависимости от его степени мы имеем или маниакальный, или меланхолический приступ с тем, что в обоих случаях обратная стадия протекает лишь в крайне умеренной степени и кратковременно. Там жег. где самоотравление возобновляется время от времени, проявляясь по силе в виде двух степеней, мы имеем двойственный психоз. Наконец, в третьих случаях мы имеем условия для постоянного самоотравления, которое колеблется лишь. в степени, давая основу для так называемого циркулярного или кругового психоза.
Очевидно, что сущность вопроса заключается не в признании или отрицании непереодических форм меланхолии и мании, а: в том, что хотя бы меланхолические и маниакальные состояния встречались на расстоянии большого периода времени или встречались совершенно изолированно, не возобновляясь второй раз в течение жизни, тем не менее они имеют между собой внутреннее родство в смысле одного и того же патогенеза, какое только, можно себе представить. Поэтому все болезненные состояния, будут ли они представлять собой простую меланхолию и манию или периодические формы того и другого состояния, или, наконец:, так называемый круговой или двойственный психоз, должны быть отнесены к одной и той же болезненной форме как отдельные ее разновидности, причем в силу этого общая болезненная форма должна получить одно общее наименование маниакально-меланхолического психоза вместо не совсем правильного названия маниакально-депрессивного психоза.
О ТЕРАПЕВТИЧЕСКОМ ЗНАЧЕНИИ
ВОСПИТАНИЯ СОЧЕТАТЕЛЬНЫХ РЕФЛЕКСОВ
ПРИ ИСТЕРИЧЕСКИХ АНЕСТЕЗИЯХ
И ПАРАЛИЧАХ
Многократные попытки исследования кожных рефлексов, уколами у истеричных нередко приводят к тому, что анестезия, бывшая в начале исследования ясно выраженной, начинает постепенно ослабевать, первоначально на месте раздражения уколами, а затем и в остальных частях пораженной области. То же наблюдается и при воспитании сочетательных двигательных рефлексов, если анестезированная область подвергается методическому электрическому раздражению для получения сочетательного двигательного рефлекса по выработанному у нас методу. Мало-помалу анестезия ослабевает и, наконец, исчезает совершенно.
Неоднократно также случалось наблюдать, что если у больных имеется истерический паралич, то воспитание сочетательных рефлексов на парализованной конечности приводит к тому,*, что вместе с появлением сочетательного рефлекса движения в парализованных областях начинают постепенно восстанавливаться и, наконец, истерический паралич излечивается более или менее совершенно.
Таким образом, вслед за воспитанием сочетательного двигательного рефлекса на парализованной конечности идет и восстановление тех движений, которые мы называем личными, или, как их обыкновенно называют, произвольными.
Эти наблюдения, пока еще не очень многочисленные, дают основание полагать, что в методике воспитания сочетательных двигательных рефлексов мы имеем одно из терапевтических средств, приводящих к излечению истерических анестезий и истерических параличей.
Обращаясь к объяснению этого факта, молено, конечно, поставить вопрос, имеется ли в данном случае внушение особого рода или же прямое растормаживание угнетенной функции. Пови-димому, следует держаться второго мнения, хотя не исключена возможность, что в этих случаях играет известную роль и вну-
43,"
■шение. Зо всяком случае, как бы ни решался этот вопрос, дело идет здесь о важном терапевтическом методе, быстро устраняющем болезненные расстройства.
Ввиду того что так называемое психическое лечение анестезии и параличей у истеричных требует, вообще говоря, особых приемов, в которых играет роль в той или иной мере искусство и опытность врача, вышеуказанный терапевтический метод, с нашей точки зрения, заслуживает особого внимания, тем более, что оно не утрачивает своего значения и там, где, благодаря тем или иным условиям, обнаруживается более или менее явное противодействие как внушению, так и другим писихотерапевтическим приемам. Так как имеются наблюдения, что глухота у истеричных быстро исчезает по мере того, как у больных воспитывается сочетательный двигательный рефлекс на звуковые раздражения, то есть все основания полагать, что и другие истерические или так называемые функциональные симптомы подобного же типа, независимо от того, будут ли они встречаться у истериков или у других невротиков, должны поддаваться лечению вышеуказанным методом.
В этих видах такого рода наблюдения должны быть продол-жены в разных направлениях, чтобы выяснить позднее значение нового терапевтического метода, что будет сделано по накоплении клинического материала.
О ПСИХО-РЕФЛЕКТОРНОЙ ТЕРАПИИ г
В то время, как рефлексология начинает пробивать себе пути в область патологии, служа к объяснению некоторых из патологических проявлений нервной системы, в числе которых мы назовем, кроме многих других, наркоманию, болезненные привычки разного рода (например, онанизм и др.), различные фобии и половые извращения, ибо эти состояния не могут быть понимаемы иначе, как в форме патологических сочетательных рефлексов 2, необходимо иметь в виду, что рефлексология открывает себе путь и в область терапии нервных болезней.
Примером могут служить те же болезненные состояния психогенного происхождения, о которых речь была выше. Психо-реф-лекторное воздействие во всех болезненных состояниях психогенного происхождения должно преследовать две цели: с одной стороны по возможности затормозить возникший при известных условиях и упрочившийся патологический сочетательный рефлекс, а с другой стороны, привить или воспитать новый сочетательный рефлекс в соответствии с требованиями здоровой жизни.
Для пояснения возьмем случай сексуального извращения, которое приходится лечить психо-рефлекторным методом. Применяя этот метод, мы пользуемся словом, как символом, который, благодаря упрочившемуся у всех людей сочетанию, замещает собою либо реальные внешние впечатления, либо определенные душевные состояния. Но, спрашивается, как при психо-рефлек-торном методе можно пользоваться словом, как символом?
Прежде всего, в случаях привычек болезненного характера мы стараемся, с одной стороны, противодействовать сложившемуся путем привычки патологическому сочетательному рефлексу, воспроизводя путем слова определенные тормозящие впечатления, указывая на несоответствие патологического сочетательного рефлекса природе организма и на происходящий отсюда моральный и физический вред здоровью и упрочивая в то же время связь их с реакцией отвращения и с необходимостью в интересах здо-
1 В. Бехтерев, Доклад в нервно-психиатрической секции Психоневрологического института, май 1915.
2 В. Бехтерев, Доклад в нервно-психиатрической секции Психоневрологического института, 1914—1915; см. также Русский врач, 1915.
28 В. М. Бехтерев
433
ровья полного воздержания от повторения патологического сочетательного μαφ-^θβ; с другой стороны, мы всемерно стараемся привить ор1анизму больного нормальный сочетательный рефлекс, для чего возбуждаем путем слова все реакции, поддерживающие этот сочетательный рефлекс, и заставляем больного по возможности осуществить его на деле, дабы сделать его привычным для организма сочетательным рефлексом, каким он и является в огромном большинстве случаев у нормальных людей.
Возьмем другой пример, представляющий случай патологического страха, или так называемые фобии. Допустим, что мы имеем фобию открытых мест. Как известно, эта фобия состоит в том, что больному навязывается мысль, что на открытом месте с ним может что-либо произойти, например, сердце его не выдержит, и он не будет в состоянии перейти открытую площадь. В таком случае психо-рефлекторное воздействие должно быть направлено на то, чтобы оживить все реакции, тормозящие развитие страха при данных условиях, и, прежде всего, путем убеждения о полном здоровье организма вообще и сердца в частности, вселить больному 'уверенность относительно невозможности какого-либо неожиданного с больным происшествия во время перехода через площадь. Этим самым уже тормозится развитие того страха и волнения, которые сочетаются у такого рода больных с одним видом большой площади.
Но помимо этого необходимо таким больным вселить уверенность относительно действительной возможности переходить, площади без всяких дурных последствий для себя и поддержать эту уверенность осуществлением спокойного перехода открытых площадей, чем упрочивается нормальный сочетательный рефлекс, свойственный всем вообще здоровым людям.
Возьмем еще пример из области наркомании. Допустим, что мы имеем наркоманию в форме привычного пьянства. В этих случаях, опять-таки, применяя психо-рефлекторный метод, мы должны стремиться к возбуждению всевозможных психических тормозов в отношении привычного злоупотребления алкоголем, связывая с последним при посредстве слова и причиняемый вред здоровью, и нравственные последствия как для самого больного, так и для его семьи, и в то же время воспроизводя перед ним материальные последствия привычного пьянства. С другой стороны, необходимо укрепить одновременно и здоровый сочетательный рефлекс воздержания путем оживления реакций, связанных с положительной стороной воздержания, как в смысле большей трудоспособности при воздержании, так и в смысле укрепления здоровья и, наконец, в смысле нравственных и материальных выгод для больного.
Таким образом и здесь, как и во всех других случаях, психорефлекторный метод лечения сводится,с одной стороны, к возбуждению, развитию и укреплению соответствующих тормозов патологических сочетательных рефлексов, а с другой стороны, к
434
возбуждению, развитию и укреплению нормальных сочетательных рефлексов, отвечающих интересам здоровья ор ганизмэ.
В заключение нужно остановиться еще на одном способе психорефлекторной терапии при наркомании и вообще при привычных ненормальных склонностях. Этот способ заключается в упрочении путем сочетания привычного наркотического яда с реакцией отвращения, вызываемой вместе с его приемом искусственными физическими приемами. Таким способом я давно уже пользовался при отучении больных от курения табаку. Для этой цели мной применяется слабый раствор arg. nitrici для полоскания горла, вследствие чего не только самое курение табаку,по даже умственное воспроизведение этого курения сопутствуется возбуждением реакции отвращения от противного впечатления, вызванного раздражением табачным дымом воспаленной слизистой оболочки носоглоточного пространства.
Естественно, что если такая реакция будет поддержана в течение продолжительного времени, то курение будет в конце концов оставлено, как акт, не только не возбуждающий привычной реакции удовлетворения, но и возбуждающий на место первой реакции отвращение.
Далее известно, что народ пользуется для лечения хронического алкоголизма в некоторых случаях не без успеха примесью к водке рвотных веществ, возбуждающих отвращение. Этот метод, применявшийся мною в отдельных случаях, требует еще научной разработки, которая была начата между прочим в противоалкогольном Институте одним из его врачей, но была затем временно прервана по не зависящим от него обстоятельствам.
Ясно, что и этот метод лечения алкоголизма основан на том же принципе, как и метод отучения от курения табаку с помощью полоскания горла раствором arg. nitrici, т. е. на принципе возбуждения, развития и упрочения сочетательного рефлекса отвращения и рвоты при одном виде спиртного напитка, возбуждавшего ранее сочетательный рефлекс удовлетворения.
28*
О СОЧЕТАТЕЛЬНО-РЕФЛЕКТОРНОЙ ТЕРАПИИ 1
Вместе с развитием учения о сочетательных рефлексах, исследуемых мною в сотрудничестве с целым рядом лиц первоначально на животных, а затем с начала текущего столетия на людях, стала выясняться особая роль искусственного воспитания сочетательных рефлексов в смысле восстановления нарушенных функций нервной системы при неврозах. Как известно, сочетательный двигательный рефлекс в постановке нашей методики получается путем многократного совмещения, вызванного электрическим раздражением двигательного рефлекса с индиферент-ным (в смысле данного рефлекса) звуковым, световым и другим раздражителем, после чего тот же двигательный эффект получается и на одни звуковые, световые и другие раздражения. И вот оказалось, что этот же прием получает терапевтическое значение при анестезиях и параличах невротического происхождения. В этом отношении еще в 1918 г. в редактируемом мною «Обозрении психиатрии, неврологии и экспериментальной психологии» я писал следующее: «Многократные попытки исследования уколами кожных рефлексов у истеричных нередко приводят к тому, что анестезия, бывшая в начале исследования ясно выраженной, начинает постепенно ослабевать, первоначально на месте раздражения уколами, а затем и в остальных частях пораженной области. То же наблюдается и при воспитании сочетательных двигательных рефлексов, если анестезированная область подвергается методическому раздражению с помощью электрического тока для получения с нее сочетательного двигательного рефлекса по выработанному у нас методу. Мало-помалу анестезия ослабевает и, наконец, исчезает совершенно.
Неоднократно также случалось наблюдать, что если у больных имеется истерический паралич, то воспитание сочетательных рефлексов на парализованной конечности приводит к тому, что вместе с появлением сочетательного рефлекса движения в парализованных областях начинают постепенно восстанавливаться и, наконец, истерический паралич излечивается более или менее совершенно.
1 Доклад на съезде по охране здоровья детей и под росткоз 16/XI 1925 г.
436
Таким образом, вслед за воспитанием сочетательного двигательного рефлекса на парализованной конечности идет и восстановление тех движений, которые мы называем личными или, как их обыкновенно называют, произвольными» 1.
С тех пор наблюдения в этом отношении проводились в том же направлении и можно было убедиться, что в методе воспитания сочетательных двигательных рефлексов мы имеем действительное лечебное средство против анестезий и параличей при неврозах.
Не менее благоприятные результаты были получены у нас и при воспитании сочетательных рефлексов на звуковые раздражения у лиц с функциональной глухотой при травматических неврозах, а также и при истерии. Вместе с многократной пробой воспитывать по выработанному у нас методу сочетательный рефлекс у оглохших от контузии постепенно, но в общем быстро, чаще всего в течение нескольких сеансов, совершенно утраченный ранее слух восстанавливался.
По показанию такого же рода больных вместе с воспитанием сочетательных рефлексов у них как бы «отворяются» уши.
Очевидно, что и истерическую слепоту можно исправлять точно таким же приемом. Во всяком случае, в методе воспитания сочетательных рефлексов мы получаем важное терапевтическое средство, дающее возможность быстро исправлять истерические параличи и анестезии кожной поверхности и специально воспринимающих органов. Очевидно, что функционально заторможенные корковые связи в этих случаях под влиянием соответствующих раздражителей и установления сочетательных связей между ними и двигательным аппаратом подвергаются растормаживанию, чем и достигается в данном случае восстановление функциональной деятельности.
Позднее методика воспитания сочетательных рефлексов существенно расширилась. Так, при работах в заведываемом мною Государственном рефлексологическом институте по изучению мозга явилась возможность, с одной стороны, получать сочетательный рефлекс путем сочетания активного движения рукой при словесном раздражителе «действуй» с одновременно производимым индиферентным раздражением, например, звуками, после чего это движение осуществляется и при одном индифе-рентном раздражении при посредстве звука.
Таким образом, мы здесь имеем настоящий сочетательный рефлекс в виде Движения руки на один звук. При этом выяснилось,что такие же сочетания пассивного движения руки со звуком не дают возможности воспитывать сочетательный рефлекс вышеуказанного характера (Дернова, Ярмоленко). Точно так же,
1 См. мою работу: «О'терапевтическом значении воспитания сочетательных рефлексов при истерических анестезиях и параличах», Обозрение психиатрии', 1917—1918, стр. 54—55, № 1—2.
4?7
■цели звуковой раздражитель совмещать с словесным раздражителем «подними» и вместе с тем испытуемый будет производить активное поднятие пальцев руки от ключа, на котором они лежат, то при повторении этих сочетаний легко образуется сочетательный рефлекс отдергивания пальцев руки на одно звуковое раздражение.
В дальнейшем оказалось возможным пользоваться сочетанием возбуждающего словесного раздражителя «подними», а также затормаживающего данное действие в смысле противодействующего приказа «не поднимай» (т. е. не поднимай пальцев) со звуком еще и в ином отношении. Допустим, что воспитан сочетательный рефлекс с помощью совмещения электрокожного и звукового раздражителя на звук «до». Как известно, вначале такой рефлекс недиференцирован, так как отдергивание пальцев руки происходит и на звук «до» и на звук «фа» и на всякий другой звук. Но вместо того, чтобы продолжать диференцировку постоянным подкреплением полученного рефлекса путем сочетания «до» с электрокожным раздражителем и оставлением без подкрепления такими сочетаниями других звуков, мы говорили одновременно со звуками «фа» и«не отнимая». У испытуемого в таком случае заторможивается рефлекс на «фа». Но вместе с тем торможение распространяется и на другие звуки: «ре», «ми» и пр., а иногда дже и на «до», и в таком случае для восстановления сочетательного рефлекса на «до» требуется новое его сочетание с электрокожным раздражением, которое с восстановлением сочетательного рефлекса может вновь распространить свое возбуждающее влияние на другие звуки, пока окончательно не установится соответствующий рефлекс на один звук «до».
С другой стороны, если мы имеем торможение воспитанного сочетательного рефлекса на данный звук и будем сочетать этот звук со словом «отними», мы тем самым растормозим сочетательный рефлекс на данный звук и вместе с тем растормозятся сочетательные рефлексы и на другие звуки. Опыты, которые были произведены у нас д-ром В. Н. Осиповой, показали, что словесный раздражитель тормозящего характера в большинстве случаев вызывает ответную реакцию быстрее, нежели словесный раздражитель возбуждающего характера и, с другой стороны, воздействие первого раздражителя приводит и к заторможению воспитанного ранее сочетательного рефлекса.
Вышеуказанный метод показывает значение сочетания ело- , весного раздражителя того или иного характера, т. е. тормозящего или возбуждающего, с рефлекторным раздражителем.
Это и дало основание применять вышеуказанный метод в целях рефлексотерапии. В Государственном рефлексологическом институте по изучению мозга этот метод получил применение против привычного воровства у детей дошкольного возраста и частью школьного возраста. Д-р В. Н. Осипова провела рефлексотерапию этого рода на 7 приютских детях в возрасте от 9 до
4 38
12 лет, из них 3 девочки и 4 мальчика, причем в этом числе имелись и настоящие клептоманы, и дети с привычным воровством. При этом с детьми, приводимыми для лечения в институт, во избежание условий внушения не велось предварительно никакой беседы по поводу их воровства, а делался только опрос имени, фамилии, возраста, школы, класса и не допускалось при этом никаких других вопросов. Самое же лечение состояло в следующем.
После установления степени возбудимости пальцев к электрическому току у ребенка воспитывается сочетательный рефлекс на звук и определяется скорость сочетательного рефлекса для выяснения сочетательно-возбудимого или сочетательно-тормозного типа. Затем производится сочетание такого минимального электрокожного раздражения, которое вызывает отдергивание пальцев со словом «не бери». Таких сочетаний производится 10 и этим дело на первый раз заканчивается. Подобные же приемы лечения повторяются приблизительно через неделю или большее или меньшее число дней. Впоследствии методика сводилась к сочетанию того же слова «не бери» с одним звонком и притом с одинаковым успехом. Уже после 2—3 таких сеансов наблюдалось улучшение, а после 6 сеансов наступило выздоровление, прослеженное 6—7 месяцев. Рецидив был лишь в одном случае, что могло зависеть от неподходящей обстановки. Некоторые же из выздоровевших детей сами с удивлением заявляли, что «влечение к воровству у них прекратилось благодаря лечению током». Далее, была сделана попытка со стороны В. Н. Осиповой применить тот же метод против детского онанизма при посредстве сочетания слова «не трогай» с электрическим током.
Дерново-Ярмоленко и Тарановой аналогичный метод сочетания электрического тока был применен у детей с ночным недержанием мочи, причем здесь с электрическим током сочетали словесный тормозной раздражитель «не мочись». И в этом, и в Другом случае Дело идет лишь о начале исследований и число наблюдений еще слишком незначительно, чтобы делать соответствующие выводы, но все же и в онанизме, и в ночном недержании мочи результаты такой терапии пока оказались вполне благоприятными. Эти исследования, конечно, будут продолжаться и тогда будет о них сказано окончательное слово.
Чтобы отдать себе отчет в значении словесного раздражителя в вышеуказанных случаях, необходимо иметь в виду, что он имеет всегда и везде замещающее значение по отношению к тому действию, символическим знаком которого служит. Так, в жизненных условиях «возьми», «не бери», «не трогай» основаны на прочной связи этих словесных раздражителей с условиями мер воздействия— поощрительных при соответствующем выполнении, противодействующих при невыполнении этого приказа. Очевидно, что в вышеуказанных случаях многократное вкоренение такого словесного раздражителя в связи с основным раздражителем,
439
возбуждающим оборонительный рефлекс, а затем и с сочетанным с ним раздражителем достаточно, чтобы возыметь соответствующее лечебное воздействие.
Естественно, возникает вопрос, не имеем ли мы Дело в последних случаях с внушением или же дело идет о прямом растор-маживании угнетенной функции в одних случаях или стимулировании активной задержки в других. Этот вопрос тем более уместен, что и в клептомании, и в онанизме, и в ночном недержании мочи я на основании своего опыта имел прекрасные результаты с помощью психотерапии отвлечением г.
Так как и само внушение с рефлексологической точки зрения следует рассматривать как растормаживание путем словесного раздражителя в одном случае (например, при устранении навязчивого влечения) и установление и упрочение связи между внешним раздражителем и действием в другом случае (например, когда делается внушение: «при виде такого-то предмета вы сделаете то-то»), то, повидимому, нельзя видеть существенного различия между вышеуказанным приемом с применением сочетательного рефлекса на словесный раздражитель и внушением, но все же в применяемом методе мы имеем более механизированный процесс сочетания высших рефлексов с рефлексогенным, или ин-диферентным раздражителем, нежели практикуемое внушение, вследствие чего не может быть и противодействия со стороны внушаемого с характером противовнушения, с другой стороны, в нем нет и следов гипнотического состояния.
Возьмем пример Тогау — лечебного приема против алкоголизма, который состоит в даче несколько раз рвотного вместе с вином и который основан на непосредственном сочетании двух раздражителей — одного с характером влечения и другого, более сильного раздражителя, с защитным характером. Можно ли говорить в этом случае о внушении? Полагаю, что нет. Другой пример: в моей практике неоднократно имелось налицо благоприятное успокаивающее действие вводимого под кожу физиологического раствора взамен привычного впрыскивания раствора морфия, что можно вернее всего объяснить установившимся путем сочетания благотворным действием самого укола на общее состояние организма, нежели внушением.
Бо всяком случае вышеуказанные лечебные приемы с характером рефлексотерапии заслуживают особого внимания ввиду того, что они дают в общем прекрасные результаты и в тех отдельных случаях, где применение гипнотического внушения и других видов психотерапии может не дать соответствующих результатов.
1 В. Бехтерев, Cber die Behandlung der krankhaften Triebe und ZwangszustSnde mit Neuerzeihung durch Ablenkungstherapie, Zschr. f. Neur. u. Psych., 1924; Гипноз, внушение и психотерапия, Вестник знания, 1911.
440
Выводы
1. Сочетательно-рефлекторная терапия, будучи близка гю своей природе к внушению, отличается от него более механизированным характером, лишающим возможности осуществлять со стороны испытуемого противовнушение.
2. Сочетательно-рефлекторная терапия, являясь новым средством лечебного воздействия, в соответствующих случаях может оказать безусловно благоприятное воздействие.
3. Особенное значение эта терапия может иметь при истерических параличах и анестезиях и при истерической глухоте.
4. При болезненных и привычных влечениях сочетательно-рефлекторная терапия, как показывали опыты над воровством у детей, дает также полезные результаты там, где внушение может не оказать соответствующего воздействия.
PSEUDOPOLIMELIA PARAESTHETICA В ФОРМЕ МНИМЫХ ЧЛЕНОВ
В 1894 г. я опубликовал работу под заглавием «Своеобразное распределение в параличе чувствительности и движения» и пр. 1. В этом случае дело шло об огнестрельномдробовом ранении в верхние отделы шейного спинного мозга на границе с продолговатым мозгом. Из всех других симптомов заболевания, между прочим, обращал на себя внимание симптом, заключающийся в следующем: «В первое время вслед за ранением больному вместе с особым чувством тяжести в правой ноге постоянно казалось, что его правая нога согнута в колене, несмотря на то, что она на самом деле вытянута; благодаря этому больной, находя для себя неудобным согнутое положение своей конечности, нередко просил окружающих ее выпрямить. Ощущение это преследовало больного даже и в том случае, когда он собственными глазами убеждался в том, что его нога вытянута, а не согнута». Одновременно с этим у больного имелся паралич движения, резкое притупление осязания и восприимчивости к давлению при утрате способности определять положение членов на правой стороне. При обсуждении этого своеобразного симптома я признавал в нем проявление галлюцинации в области определения положения членов и мог сравнить его с тем состоянием, которое испытывают ампутированные под влиянием раздражения, возникающего в перерезанных нервах конечностей, когда им кажется присутствие ампутированной конечности.
В позднейшей работе П. А. Останкова (из моей клиники)2 вышеуказанный симптом получил наименование мнимых или ложных конечностей pseudomelia paraesthetica. В этом случае, как и в моем, «конечность чувствуется в согнутом положении», и так же в ней ощущается тяжесть, «наконец, в обоих случаях коррекция зрения нисколько не устраняет тягостные для больного ощущения». Необходимо иметь в виду еще и постоянство парестезии, которая держится без перемен в течение нескольких недель. Случай П. А. Останкова, находившийся под наблюде-
1 См. Ц. Бехтерев, Неврологический вестник, 1894, стр. 1 — 42.
2 П. А. Останков, Обозрение психиатрии, 1904, №9, стр. 685 и след.
442
нием совместно с другим моим ассистентом А. И. Карпинским, по локализации поражения также близок к моему, так как в нем была поражена та же верхняя область шейного мозга с участием прилегающей к ней области продолговатого мозга (судороги в губах, по временам одышка, учащение пульса, безгласие и пр.).
В этом случае мы имеем следующие симптомы: «полную вялую параплегию верхних и нижних конечностей с полным параличом в них всех видов чувствительности, задержку мочеотделения и дефекации, постоянную тоническую судорогу в шейных мышцах, судорожные подергивания в мышцах губ, легкий парез небной занавески, тихий голос, приступы одышки, слабость движений диафрагмы, отсутствие коленных рефлексов, симптом Бабинского, резко выраженный холодовый рефлекс, отсутствие кожных рефлексов, за исключением подошвенного, искривление позвоночника на уровне шейных и верхних грудных позвонков и, кроме всех этих симптомов, самый мучительный для больного — это чувство ложных конечностей». Только что перечисленный сим-птомокомплекс, толчкообразное развитие заболевания с некоторыми ремиссиями и быстрое его затем прогресирование, одновременное специфическое поражение костной системы (периоститы, распознанные специалистами) дают нам право на распознавание в данном случае люэтического менингомиэлита, локализующегося в шейном отделе спинного мозга с участием в процессе нижнего отдела продолговатого мозга (meningomielitis cervica-lis luetica et irritatio medullae oblongatae).
Интересно, что в этом случае симптом имел двусторонний характер, ибо он обнаруживался в обеих руках и обеих ногах. Автор, между прочим, отмечает, что периферические раздражения, каковы перемена положения конечности, уколы, тепло и холод, не изменяют тягостной парестезии ложных конечностей, происхождение же этого симптома он относит к раздражениям центрального отрезка проводников чувствительности и мышечного чувства.
Из первоначально опубликованного мною случая с дробовым поражением спинного мозга и только что рассмотренного случая видно, что симптом этот может быть наблюдаем при поражениях верхних отделов спинного мозга. В другом позднее опубликованном мною случае было обнаружено размягчение в области п. lenticularis '. Следовательно, вышеуказанный симптом может быть и церебрального происхождения. Приведем вкратце описание этого случая. Здесь дело шло о хроническом алкоголике 38 лет, который с определенного времени (с 1883 г.) стал испытывать галлюцинации слуха с последующим образованием идей преследования. В 1884 г. около недели больной помещался в Саратовской психиатрической лечебнице. Затем 28 января 1887 г.
1 В. Бехтерев, Обозрение психиатрии, 1904, № 12, стр. 89S
И C.'KYI.
•МЗ
он поступил в Казанскую окружную лечебницу, где я его и наблюдал вместе с врагами лечебницы. Больной обнаруживал слуховые галлюцинации с враждебным для больного характером. С июля 1887 г. у больного обнаружены явления туберкулеза. 29 февраля больной внезапо упал с кровати, после чего у него обнаружен парез правого верхнего века, расширение левого зрачка, отклонение языка влево, затрудненный выговор слов при общем упадке сил и нарушенной ориентировке. 2 марта отмечено судорожное тоническое напряжение мышц обеих левых конечностей. На другой день паралич тех же конечностей. В начале открытия исследование показало при общей худобе заметно выраженную атрофию мышц левых конечностей и лопаточно-плечевой области. Резко выраженный паралич всей левой половины тела. Левая рука слегка полусогнута, но насильно может быть разогнута. Реакция мышц на токи слева ослаблена. Коленный рефлекс слева передается на правую конечность. В левой верхней конечности мышечное чувство (определение положения) совершенно утрачено, в левой ноге ослаблено. Другие виды восприимчивости, как болевая, осязательная и температурная, сохранены. Обращает на себя внимание в периоде развития паралича у больного упорное ложное ощущение двигающейся руки слева. Больной, не имея возможности двигать своей левой рукой, чувствует, однако, присутствие двигающейся левой верхней конечности, которая иногда закидывается за спину и как бы сдавлена лежащим на ней туловищем, в другой раз она лежит на его груди или животе. Больной вполне сознает, что это чувство ложное, так как знает, что его левая рука совершенно парализована, тем не менее не может отделаться от своего ощущения даже и в том случае, когда смотрит на свою неподвижно лежащую парализованную левую верхнюю конечность.
При вскрытии мозга обнаружены: помутнение мягких оболочек, местами сращение с мозговым веществом. Сосуды основания без существенных изменений. В правом полушарии размягчение в передней части бокового желудочка серовато-желтого цвета. Размягчению подверглась большая часть п. lenticularis за исключением незначительного переднего его отдела с прилежащей частью белого вещества. Corpus caudatiim и весь thalamus не поражены. В спинном мозгу на всем протяжении заметное уменьшение размеров левого переднего рога. При микроскопическом исследовании спинного мозга обнаружена атрофия клеток переднего левого рога. Отмечается также перерождение и атрофия левого бокового столба.
Не касаясь других явлений данного случая, отметим, что своеобразный симптом присутствия иллюзорной движущейся верхней конечности при параличе руки наблюдался в данном случае при утрате лишь одного мышечного чувства или способности определять положение членов, вследствие чего этот симптом здесь должен быть отнесен к церебральному поражению об-
444
ласти п. lenticularis и соседнего белого вещества, ибо атрофиче-ские явления переднего рога с перерождением в левом боковом столбу должны быть признаны явлениями вторичными.
Во всех трех вышеуказанных случаях дело идет о своеобразном симптоме, который в свое время был назван парестетиче-ским явлением с характером присутствия ложных конечностей, или парестетической псевдомелией. Не следует забывать, что эта парестезия в сущности представляет собой особый вид «телесных» иллюзий в форме присутствия, кроме членов своего тела, еще других иллюзорных членов, находящихся в покое или движении.
Из приведенных наблюдений явствует, что такие явления связаны с нарушением способности определять положение членов в пространстве. Однако от случаев tabes 'а, при котором мы имеем такое же нарушение способности определять положение членов, вышеуказанные случаи отличаются тем, что здесь дело идет не об •одном только неузнавании положения своих членов, а о мнимом присутствии других иллюзорных членов, наряду с имеющимися парализованными или ампутированными. Это обстоятельство, как и упорство этих явлений, не исчезающих и при контроле зрением, показывает, что дело идет здесь о явлениях, обусловленных раздражением центростремительных проводников при существующем нарушении их проводимости наподобие, например, эксцентрических болей, развивающихся от раздражения проводников анестезированной области. И в самом деле в двух из вышеуказанных случаев имелись явления, указывающие на состояние раздражения в месте поражения.
Последний случай, между прочим, говорит за то, что для появления этого симптома не имеет существенного значения поражение других центростремительных проводников, кроме тех, которые служат для определения положения членов. Отсюда приходится допустить, что поражения с характером раздражения только что указанных проводников на всем протяжении от периферии до центров головного мозга включительно сопровождаются в некоторых случаях, согласно закону эксцентрической проекции, парестезией или галлюцинацией в виде иллюзорных конечностей.
Небезынтересным представляется тот факт, что в отдельных случаях дело идет о парестезии такого же рода, но не одной только, а о нескольких ложных членах тела, включая в том числе и голову. Вот одно из таких наблюдений в случае левосторонней гемиплегии, зависящей от эмболии ветвей a. fossae Sylvii.
Больной банковский служащий, 43 лет, происходит из здоровой семьи. Давно страдал недостаточностью клапанов аорты, характеризующейся резкими систолическими шумами. В остальном представлялся здоровым. Около начала октября 1924 г. он, занимаясь в банке, без каких-либо предшествующих условий почувствовал себя дурно и в течение 2—3 часов у него без явлений апоплексии парализовалась левая рука, нога и лицо. Он был доставлен в карете скорой помощи в Максимилиа-новскую лечебницу, куда я был приглашен в качестве консультанта осмотреть больного 3 ноября 1924 г. В первое время после инсульта стала
445
нарастать температура тела, поднявшаяся до 40°. Поясничным проколом обнаружено повышенное давление жидкости. При исследовании оказалась затылочная контрактура и симптом Кернига. Левосторонняя гсми-плегия, причем левый глаз не закрывается при открытом правом глазе, тогда как правый глаз закрывается свободно при открытом левом глазе. Нижняя часть левой стороны лишь слегка опущена. Носогубная складка сглажена. Язык высовывается почти прямо. Левая щека при активных усилиях влево не оттягивается. Левая рука не обнаруживает никаких вообще движений; левая нога может лишь слегка сгибаться, других же движений в ней не обнаруживается. Пассивно левые рука и нога сгибаются без сопротивления. Коленный рефлекс слева не повышен, намек на клонус и едва заметный Бабинский. Кожная восприимчивость слева понижена. Способность определять положение членов на левой стороне также понижена. Но вот что заявляет больной о своем состоянии. Бременами он испытывает, что у него на левой стороне две руки и от двух до шести нижних конечностей и три головы. Если он при этом двигает левой рукой и левой ногой или они пассивно смещаются, то он испытывает смещение только одной руки или ноги, о остальные нижние конечности лежат на месте. То же происходит и с головой при ее смещении, одна голова двигается, а другие остаются па месте. Этот симптом, наряду с параличом у больного, оказался очень стойким, продолжаясь в течение многих месяцев.
На основании данных исследования в этом случае приходится признать поражение правого полушария с характером эмболии центральных ветвей сильвиевой артерии и с последующим размягчением, причем в поражение вовлечены внепирамидные системы, а в умеренной степени и пирамидные пути ввиду вялого характера паралича со слабыми намеками на симптом Бабинского и на клонус стопы.
В данном наблюдении заслуживает внимания то, что парестезия в виде мнимых членов обнаруживалась не только в отношении конечностей, но и головы, чего в прежде опубликованных случаях не наблюдалось. С другой стороны, особый интерес представляет то обстоятельство, что здесь дело шло о парестезии не в виде одного только иллюзорного члена, кроме парализованного, но в виде нескольких членов: иллюзорных три головы, две руки и в разное время от двух до шести нижних конечностей. В этом, как и в ранее опубликованных случаях, мнимые конечности находились в покойном положении и это покойное положение оставалось таким же и при смещении существующего парализованного члена. Между тем в иных случаях, как это было в одном нашем наблюдении (третьем из приведенных), мнимые конечности могут казаться находящимися в движении. Между прочим, это явление наблюдается иногда и при периферических поражениях. В одном из таких случаев, благодаря раздражению периферической культи, больная мучилась тем, что будто бы ампутированная в плечевом суставе верхняя ее конечность вращалась в этом суставе с необычайной быстротой, что чрезвычайно тяготило больную. Отсюда представляется необходимым различать две разновидности псевдомелии, или псевдополимелии, как и следовало бы назвать случаи со многими мнимыми конечностями: 1) пассивную, когда мнимые конечности остаются в пс-ксе, и 2) активную, когда мнимые конечности находятся в дви-
4-16
жении. При этом парестезия последнего рода представляется всегда более тягостной по сравнению с первой.
В .заключение заметим, что парестетическая псевдомелия, или псевдополимелия, необязательно наблюдается при существовании паралича, ибо в одном случае при явлениях алкоголизма с слуховыми галлюцинациями без резких параличных явлений я имел возможность наблюдать одностороннюю псевдомелию в крайне ярко выраженной форме. Здесь, таким образом, дело шло уже о самостоятельной «телесной» галлюцинации, что можно наблюдать в редких случаях и у других лично (душевно) больных.
Что касается патогенеза этого, если мы обратимся к выяснению природы рассматриваемого симптома, то, очевидно, необходимо иметь в виду состояние раздражения проводников, передающих к центрам мышечную восприимчивость и дающих возможность определять положение членов в пространстве. В заключение заметим, что по своему характеру данный своеобразный симптом имеет некоторую аналогию с кожной полиэстезией и монокулярной диплопией и полиопией.
Общие выводы работы можно формулировать следующим образом:
1. Псевдополимелия в виде «телесных» иллюзий или галлюцинаций, характеризующихся присутствием иллюзорных членов тела (руки, ноги, головы), может наблюдаться при поражениях на всем пути центростремительных проводников, служащих к определению положения членов, начиная от периферии до мозговых центров.
2. В основе этого симптома лежит раздражение упомянутых проводников, обусловленное тем или иным местным их поражением.
3. Рассматриваемый симптом может состоять не только в виде одного иллюзорного члена, но и нескольких иллюзорных членов (например, 2 головы, 5—6 ног и т. п.).
4. Иллюзорные члены могут оставаться в одних случаях в покое, в других случаях могут быть в движении, вследствие чего мы можем различать пассивную и активную псевдополимелию.
ПРИМЕЧАНИЯ
К СТАТЬЯМ В. М. БЕХТЕРЕВА, ПОМЕЩЕННЫМ В СБОРНИКЕ 1
1 Авторы примечаний—сотрудники Института им. В. М. Бехтерев Г. Б. Абрамович, Г. IO. Белицкий, В. И. Буторин, Р. А. Заченицки: В. Н. Мясищев, С. Ф. Семенов,
29 Б. М. Бехтерев
I. АНАТОМИЧЕСКИЙ РАЗДЕЛ
1. «Экспериментальные исследования относительно перекреста ro-локон зрительного нерва в chiasma ηη. opticorum» (Еженедельная клиническая газета, 1883, № 2).
Вопрос о неполном перекресте волокон зрительного нерва в chiasma у высших животных и человека был спорным. Сторонники неполного перекреста основывали свои заключения на клинических наблюдениях и неирогнстологических исследованиях. В. М. Бехтерев впервые методом перерезки chiasma показал, что у собак нет полного перекреста зрительных нервов. Таким образом, приоритет экспериментального подтверждения неполного перекреста волокон зрительного нерва в chiasma принадлежит В. М. Бехтереву. Дальнейшие исследовании Пинеса, Моиа-кова, Миньковского подтвердили данные В. М. Бехтерева.
2. «Об одной до сих пор неизвестной связи между главной оливой и головным мозгом» (Neurologisches Zentralblatt, 1885, № 9).
В этой статье В. М. Бехтерев впервые у новорожденного месячного возраста описывает имеющий большое физиологическое значение центральный пучок покрышки, которой связывает большую оливу с вышележащими отделами мозга. До В. М. Бехтерева зарубежными авторами этот пучок ошибочно считался продолжением бокового канатика (Штиллинг) или пучком покрышки из задней комнссуры (Вернике) (перевод статьи сделан В. И. Френкелем).
3. «О соединениях верхних олнв и об их вероятной физиологической роли» (Врач, 1885, № 32).
В этой работе В. М. Бехтерев подробно описывает связи верхней оливы с задним четверохолмием (через боковую петлю), мозжечком, ядром отводящего нерва, передним ядром слухового нерва, а также со спинным мозгом. В ней также приводятся некоторые соображения автора в отношении вероятной физиологической роли верхних олив.
4. «О двух пучках, входящих в состав внутреннего отдела задней ножки мозжечка, н о развитии волокон слухового нерва» (Врач, 1885,
•№ 25).
В. М. Бехтереву принадлежит приоритет в открытии особого ядра нервных клеток в головном мозгу. Он впервые описал мелкоклеточное скопление дорзальнее ядра Деитерса, имеющее отношение к корешку (вестибулярному) слухового нерва. Это скопление клеток в дальнейшем получило название «ядра Бехтерева»; оно лежит кнаружи от угла четвертого желудочка и служит местом окончания преддверного нерва.
5. «О задних корешках, месте их окончания в сером веществе спинного мозга и об их центральном продолжении» (Медицинское обозрение, т. 29; Вестник клинической и судебной психиатрии, 1887, в. 1).
В. М. Бехтереву принадлежит приоритет описания распределения в сером и белом веществе центральной нервной системы задних корешков спинного мозга. Он показал, что задние корешки состоят нз неоднородных волокон, получающих миэлнновый покров в разное время, имеющих различный калибр и вступающих в соотношение с различными аппа-
2д*
4S1
ратами спинного мозга. Так, внутренний, ранее развивающийся пучок состоит из волокон толстого калибра, наружный, позднее развивающийся пучок состоит из тонких волокон.
6. «К вопросу о наружных сочетательных волокнах мозговой коры» (Медицинское обозрение, 1891, т. 36, № 22, стр. 926—928).
В статье В. М. Бехтерев впервые описывает пучок волокон, представляющий собою наружный ассоциационный слой мозговой коры. Этот слой получил в дальнейшем название «бехтеревской полоски».
Кезом позднее В. М. Бехтерева среди тонких волокон мозговой коры был описан особый пучок волокон, который он необоснованно назвал «наружным мейнертовским ассоциационным слоем». Этот пучок по своему расположению аналогичен бехтеревской полоске.
7. «К вопросу о мозговых полосках продолговатого мозга» (Медицинское обозрение, 1892, т. 37, № 5).
В статье В. М. Бехтерев подробно излагает анатомические данные о мозговых полосках продолговатого мозга, независимость которых от слуховых нервов была нм впервые доказана еще в 1887 г. Кроме того. в этой работе виден приоритет В. М. Бехтерева в открытии того факта, что передний корешок слухового нерва происходит из преддверной части (г. vestibularis) и что позднее развивающийся задний корешок происходит из улитковой части (г. cochlearis) слухового нерва.
8. «Об «олнвчатом» пучке шейной части спинного мозга» (Неврологический вестник, 1894, т. 2, в. 2).
В настоящей статьг В. М. Бехтерев описывает выделенный им по методу развития особый, поздно развивающийся пучок, принадлежащий главным образом шейной части спинного мозга и отчасти нижнему отделу продолговатого мозга и имеющий ближайшее отношение к большой оливе. Этот пучок вместе с описанным В. М. Бехтеревым центральным пучком покрышки, повидимому, составляет единую систему волокон, прерываемую на своем пути в нижней оливе. В. М. Бехтерев называл этот пучок «оливчатым пучком шейной части спииного мозга», а не «треугольным пучком», как его обозначил Гельвег. Описание этой системы произведено В. М. Бехтеревым совершенно независимо от Гельвега, задолго до него, и сделано более точно, чем Гельвегом.
9. «Об ядрах глазодвигательных нервов (ηη. oculomotorii, abducen-tis и-trochlearis) и о связи их друг с другом» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 7).
Несмотря на то, что работа опубликована более 50 лет назад, приведенные в ней данные соответствуют современным представлениям об ядрах глазодвигательных нервов и о связях их друг с другом, дополняя их материалами, полученными эибриогенетнческим методом исследования. На основании исследования мозгов у зародышей и у новорожденных В. М. Бехтерев устанавливает наличие четырех ядер глазодвигательного нерва, а также связи их с соседними образованиями (задним продольным пучком, передним двухолмием), ндром отводящего нерва. Им установлена сочетательная система волокон между ядрами глазодвигательного иерва.
10. «Об особом внутреннем пучке боковых столбов» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 4).
В этой статье В. М. Бехтерев приводит данные, касающиеся впервые открытого нм по методу развития в задней части пограничного слоя особого пучка, который он обозначает как внутренний пучок боковых столбов спинного мозга.
11. «Об особом ядре сетевидного образования на уровне верхних отделов варолиева моста» (Неврологический вестннк, 1898, т. VI, в. 2).
В статье В. М. Бехтерев, помимо описанных им ранее четырех гнезд серого вещества сетевидного образования, впервые описывает верхнее центральное ядро сетевидного образования, наличие которого он отмечает у кошки, собаки, а также у человека.
452
12. «О развитии клеток мозговой коры у человека» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 8).
Работа показывает, что по вопросу о происхождении нервных клеток мозговой коры В. М. Бехтерев не придерживался господствовавшей в то время механистической концепции Вирхова о происхождении клеток только из клеток и считал, что первоначальному образованию клеток мозговой коры предшествует рост особых волокон илн нитей, которые делают изгибы вокруг зародышевых телец, составляя остов будущей клетки; вещество волокон (нли нитей) служит для образования клеточной протоплазмы и ее отростков, в то время как зародышевые гельца служат для образования ядер. Данное исследование близко подходит к концепции О. Б. Лепешинской о развитии клеток из живого вещества.
II. ФИЗИОЛОГИЧЕСКИЙ РАЗДЕЛ
Работы В. М. Бехтерева, посвященные выяснению зависимости самых различных функций организма от деятельности головного мозга, были выполнены в допавловский период физиологии. Отсюда вытекают взгляды В. М. Бехтерева на проблему локализации функций. Позиции, занятые В. М. Бехтеревым, могут быть лучше всего охарактеризованы его же словами. В 1887 г. он писал: «...я вовсе не причисляю себя к числу авторов, которые смотрят на кору как на мозаику, состоящую из отдельных кусков различной окраски. Кора мозга, быть может, и уподобляется карте, разрисованной различными красками по отдельным участкам, но так, что соседние краски, понятно, смешиваются между собой и при этом, быть может, на этой карте не существует ни одного участка, покрытого одним цветом, а не смешанного из множества красок» *. Мы видим, что, хотя В. М. Бехтерев и не разделял узко локалисти-ческой точки зрения, но, как бы ни была сложна мозаика распределения функций на поверхности головного мозга, она представлялась ему не в том динамическом плане, который вытекает из принципа временных связен.
Разумеется, что такого рода факты могли получить новое освещение только после того, как И. П. Павловым был установлен принцип временной связи в работе коры головного мозга, чем и была заложена основа современного функционально-динамического понимания проблемы локализации.
В этой же связи следует указать, что в ряде работ В. М. Бехтерева, написанных до установления И. П. Павловым принципа временной связи, можно встретиться с непоследовательными формулировками, в которых психическое представлено в отрыве от физиологического.
Только учение о высшей нервной деятельности дало естественно-научную основу последовательно материалистическому пониманию психики как процесса отражения объективной действительности.
В своем отзыве от 20 ноября 1908 г. о книге В. М. Бехтерева «Основы учения о функциях мозга» И. П. Павлов дает ей высокую оценку, но в то же время отмечает, что «в книге нмеетсн немало и недостатков». Он пишете «недостаточной точности и определенности» изложения автора, о том, что «некоторые теоретические представления автора в значительной степени произвольны», что «примирение имеющихся в науке противоречий часто носит словесный характер», «постоянно смешиваются психологическая и физиологическая точки зрения», самым же существенным недостатком книги И. П. Павлов считал «недостаточное критическое, недостаточное объективное отношение к исследованиям собственной лаборатории автора».
1. «Об отправлении зрительных бугров» (Врач, 1883, № 4 и 5).
Работа опубликована в 1883 г. и содержит в сокращенном виде основные факты и выводы, которые впоследствии были изложены В. М. Бехтеревым подробнее в ряде статей, не вошедших в настоящее нз-
1 В. М. Бехтерев, Физиология двигательной области мозговой коры, Архив психиатрии и неврологии, 1887, т. IX, 2—3.
454
дание ввиду их большого объема. В. М. Бехтерев на основании большого числа экспериментов на животных и клинических наблюдений на человеке впервые создал представление, согласно которому область зрительных бугров имеет непосредственное отношение к совокупности двигательных и вегетативных реакций, выражающих эмоциональное состояние животных и человека. Таким образом, В. М. Бехтерев намного опередил исследования Карплюса и Крейдля, которым неправильно приписывался приоритет в этой области.
В 80-х годах прошлого столетия В. М. Бехтерев начал разработку вопроса о функциональных связях между высшими отделами центральной нервной системы и организмом, в частности, между корой головного мозга и внутренними органами.
2. «Об иннервации и мозговых центрах слезоотделения» (Медицинское обозрение, 1891, т. 35, № 12).
3. «О влиянии коры большого мозга на акты глотания и на дыхание» (Неврологический вестник, 1894, т. 2, в. 2).
4. «К вопросу о влиянии мозговой коры и зрительных бугров на глотание» (Неврологический вестник, 1894, т. 2, в. 2).
В настоящем сборнике печатается часть статей В. М. Бехтерева, посвященных этим вопросам 1.
5. «О влиянии мозговой коры человека на сердцебиение, давление крови и дыхание» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 11).
6. «Влияние коры головного мозга на половые органы, предстательную и молочные железы» (Archiv fur Anatomie und Physiologic Physi-ologische Abteilung, 1905) (перевод статьи сделан Н. Э. Гранстрем и А. М. Свядощ).
До Бехтерева соответствующая литература была представлена единичными работами (В. Я. Данилевский, 1874, и др.). В систематических экспериментах В. М. Бехтерева на теплокровных животных, преимущественно собаках, был использован метод электрического раздражения коры и других центральных областей головного мозга в сочетании с наблюдением за двигательными или секреторными ответами на эти раздражения со стороны внутренних органов. В этих условиях было убедительно показано, что при раздражении определенных пунктов головного мозга функция исследуемого органа усиливается, в то время как после раздражения других участков коры работа органов, напротив, угнетается. Этим впервые с несомненностью было установлено, что в определенных участках коры головного мозга, главным образом в области сигмовидной извилины, имеется представительство для большинства внутренних органов.
Значение вышеприведенных работ несомненно не только для того времени, когда они были выполнены, т. е. для конца прошлого и начала текущего столетия, когда правомерность вопроса о корковой регуляции вегетативных функций казалась вообще сомнительной и вызывала споры.
После того как И. П. Павлов развил представление о 'временных связях как важнейшем принципе в работе высших отделов центральной нервной системы, В. М. Бехтерев стал исследовать реакции животного с удаленными участками коры головного мозга на ряд естественных условных раздражителей, с помощью этих наблюдений удалось получить дополнительные данные, свидетельствующие об отношении этих участков к ряду функций, например, к желудочному сокоотделению (Никитин, 1906), функциям половых органов (В. М. Бехтерев и Л. М. Пуссеп, 1906), дыханию (Жуковский, 1908).
Однако только исследования М. К. Петровой, К. М. Быкова и М. А. Усневича с сотрудниками, посвященные изучению кортико-вис-
1 Статьи В. М. Бехтерева, написанные совместно с проф. Н. А. Ми-славским и вошедшие в «Избранные произведения» Н. А. Миславского (Медгиз, 1952), в настоящий сборник не включены.
455
неральпых связей в норме и патологии обеспечили глубокое и многосто» роннее развитие павловских идей о влиянии коры полушарий на все процессы, происходящие в теле.
7. «О значении органов равновесия в отношении развития наших представлений о пространстве» (Неврологический вестннк, т. III, в. 4,1895).
Серия работ В. М. Бехтерева по вопросу о функциях органов равновесия и значении их для развития пространственных представлений, начавших публиковаться с 1882 г., имеет интерес не только исторический, несмотря на то, что современные представления по вопросу о локализации отдельных аппаратов системы равновесия коренным образом изменились.
Идеи В. М. Бехтерева о центральной саморегуляции систем, сигнализирующих о положении тела в пространстве и рефлекторных ответах со стороны органов движения, способствующих поддержанию равновесия и исправлению его нарушений, предвосхищают развившееся позже за рубежом учение о тонических рефлексах и рефлексах положения. Первые шаги в этой области были сделаны не Шеррипгтоном, Магнусом и др., а В. М. Бехтеревым.
Большое теоретическое значение имеют положения В. М. Бехтерева о роли органов равновесия в развитии представлений о пространстве, о механизмах пространственной локализации ощущений и нарушениях этих механизмов. В. М. Бехтерев признавал, что представления и понятия о пространстве образуются на основе наличия соответствующих апа-томо-физиологических аппаратов в процессе отражения объективной действительности.
Большой принципиальный интерес представляет критика Маха В. М. Бехтеревым. Здесь В. М. Бехтерев не смог раскрыть методологическую порочность и реакционность представлений Маха, однако он понимал ошибочность концепции Маха, который в соответствии со своими идеалистическими взглядами считал, что ощущение положения тела в пространстве есть имманентная функция организма.
Вместе с тем нужно признать неправильными также психологические формулировки В. М. Бехтерева, например, «восприятие ощущений» или «пространство, созданное ощущениями положений тела и его членов».
8. «О так называемом судорожном центре и о центре передвижения тела на уровне варолиева моста» (Неврологический вестник, 1896, т. 4, в. 4).
В статье излагаются результаты ряда экспериментов, выполненных в лаборатории В. М. Бехтерева с целью выяснить механизм судорожного приступа, развивающегося после механического повреждения варолиева •моста. Эти опыты опровергли существующее мнение о наличии судорожного центра в названной области, поскольку при этом была установлена роль коры головного мозга в развитии судорожного припадка. Важно вместе с тем отметить, что произведенные по ходу работы перерезки мозгового ствола ща различных его уровнях дали В. М. Бехтереву возможность указать на ту смену соотношений тонических и клонических компонентов судорожного состояния мышц всего тела животного, которая позже стала предметом наблюдения н послужила исходным пунктом для исследований децеребрационной ригидности.
9. «О возбудимости мозговой коры у новорожденных животных» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 7).
Исследования В. М. Бехтерева о возбудимости различных отделов головного мозга у новорожденных животных изложены в ряде статей, написанных в 80-х и 90-х годах прошлого столетия. Настоящая статья является наиболее поздним сообщением результатов этих исследований.
В. М. Бехтерев является одним из первых исследователей вопроса о развитии функции высших отделов центральной нервной системы в онтогенезе. В. М. Бехтеревым выявлен на ранних этапах постпатального
4,-)С>
периода диффузный и замедленный характер корковой деятельности, который сочетается с преобладанием тонической иннервации над тетани-ческой. Способность же воспроизводить более точно локализированные и координированные ответные реакции является достижением последующего периода индивидуального развития и определенным образом связана с деятельностью органов чувств.
В. М. Бехтерев признавал значение факторов внешней среды и, в частности, роль зрительной рецепции в организации и перестройке высших отделов центральной нервной системы на более высокий уровень конкретной и диференцированной деятельности. Он опровергает утверждение некоторых исследователей (Зольтман, 1876) относительно того, что между сроками включения зрительной рецепции н проявлением первых признаков возбудимости коры головного мозга существует прямая хронологическая связь. Здесь же в опытах с раздражением различных отделов головного мозга он признает зависимость возбудимости этих отделов мозга от сроков развития процесса миэлинизации нервных волокон.
10. «О репродуктивной и сочетательной реакции в движениях» (Обозрение психиатрии, неврологии и экспериментальной психологии,
1908, № 7).
В данной статье В. М. Бехтерев устанавливает, что сам по себе ритм, в котором работает какой-либо центр, может играть роль условного раздражителя по отношению к другому центру, вовлекая его в деятельность в том же ритме. Такого рода зависимости обозначены В. М. Бехтеревым как репродуктивные реакции. Эти исследования обогащают фактическим материалом учение о высшей нервной деятельности.
11. «Исследование функций коры с помощью естественных сочетательных рефлексов. Значение этого метода по отношению к центрам внутренних органов и различных секреций» (Обозрение психиатрии, непро логии и экспериментальной психологии, 1908, № 8).
В данной работе подводится итог ряда исследований В. М. Бехтерева преимущественно о корковой регуляции работы внутренних органов. Эта работа представляет одну из первых попыток осветить нервный механизм деятельности внутренних органов с позиций учения об условных рефлексах. В. М. Бехтерев пользуется термином «сочетательный рефлекс», хотя более точен термин И. П. Павлова — условный рефлекс. В полном объеме освещение этих вопросов и значение их дано в работах К. М. Быкова и его сотрудников о взаимосвязях коры головного мозг;] н внутренних органов.
Заслуживает внимания приводимый в работе, но еще не вполне ясный факт сохранности рефлексов в ответ на вкусовые и обонятельные раздражения при утрате их в ответ на. зрительные и слуховые после оперативных вмешательств на коре головного мозга.
III. РАЗДЕЛ НЕВРОПАТОЛОГИИ
1. «Наблюдения трофических изменений кожи в зависимости от расстройств нервной системы» (Еженедельная клиническая газета, 1881, № 27, 28).
В. М. Бехтерев в этой статье один из первых в русской литературе осветил вопрос о связи трофических расстройств кожи (сыпи, аномалии пигментации, поседение и выпадение волос) с изменениями нервной системы. Автор подчеркивает значение нервной системы в развитии нарушений кожной трофики. Автор наблюдал изменения кожи типа красного лишая, имеющего распространение по ходу кожных нервов. Ненормальную пигментацию покровов (гипертрофию и атрофию кожного пигмента) В. М. Бехтерев рассматривает как трофоневроз. Преждевременное поседение и выпадение волос также может зависеть от изменений нервной системы. Данное исследование В. М. Бехтерева представляет одно из первых приложений принципа нервизма в дерматологии.
2. «Одеревенелость позвоночника с искривлением его как особая форма заболевания» (Врач, 1892, № 36).
В этой статье впервые дается В. М. Бехтеревым клиническое описание одеревенелости позвоночника как особой формы заболевания, получившего в дальнейшем наименование «бехтеревской болезни». Болезнь В. М. Бехтерева клинически и патологоанатомически существенно отличается от формы Штрюмпель-Мари, описанной указанными авторами позже.
3. «Об анкилозирующем воспалении позвоночника и больших суставов конечностей» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 7—8).
После описания В. М. Бехтеревым особой формы заболевания одеревенелости позвоночника Штрюмпель и Мари опубликовали свои сходные наблюдения. В. М. Бехтерев в данной статье приводит диференциаль-ную диагностику наблюдений Штрюмпеля и Мари, представляющих собой хроническое анкилозирующее воспаление больших суставов и позвоночного столба.от описанной им до того формы. В. М. Бехтерев всегда отстаивал нозологическую самостоятельность выделенной им формы заболевания.
4. «О звуковой проводимости прн постукивании костей черепа и позвоночника, как методе исследования нервных болезней» (Неврологический вестник, 1894, т. 2, в. 1).
В этой статье В. М. Бехтерев приводит данные исследования проводимости звука костями черепа. Сравнением звукопроводимости симметричных и соседних областей черепа (выслушивание при помощи стетоскопа) выявляется заглушение тона звучащего камертона как признак поражения черепных костей и подлежащих областей мозга. В отношении позвоночника, кроме этого метода, автор дает описание применения глубокой перкуссии позвоночника. Исследование проводимости звука костями черепа и глубокая перкуссия позвоночника представляют большой интерес, в частности, для диагностики новообразований мозга. Этот метод недостаточно широко использован в диагностической практике в значительной степени вследствие больших требований к слуху врача. Однако
458
современная техника дает широкие возможности применения этого метода с объективной регистрацией звукопроводимости.
5. «О значении совместного употребления бромидов н Adonis ver-nalis при падучей» (Неврологический вестник, 1894, в. 3).
В. М. Бехтерев рекомендует для лечения эпилепсии комбинированное употребление бромидов и Adonis verflalis, от применения которых он всегда получал хороший эффект. Введенная Бехтеревым лекарственная смесь в настоящее время широко применяется для лечения неврозов и получила название бехтеревской микстуры.
6. «О хореической падучей» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 9).
В. М. Бехтерев описывает особый вид падучей, которую он назвал «хореической или плясковой падучей», так как при ней отмечается сочетание хореи в виде судорожных подергиваний в разных частях тела с падучей, причем хорея не только осложняет падучую, но стоит с ней в причинной связи»
Вопросы патогенеза этой формы эпилепсии, роли наследственности взаимоотношений между истерией и хореей в настоящее время освещаются иначе, чем это представлено В. М. Бехтеревым.
7. «О малоизвестных ранних симптомах спинной сухотки» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 9).
8. М. Бехтерев впервые обнаружил, что сочетание усиления кожных (особенно брюшиых и надчревных) и угасания сухожильных рефлексов нижней половины тела (коленного и ахиллова), равно как и безболезненность к давлению мышц, особенно нижних конечностей (в частности, икроножных), наряду с безболезненностью п. tibialis, являются симптомами, характерными для ранних периодов спинной сухотки. Симптомы, описанные в данной статье, существенны для диагностики сухотки спинного мозга.
8. «Апоплектическая гемитоиия» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 7—8).
На основании анализа клинических наблюдений В. М. Бехтеревым впервые выделен описываемый в данной статье особый синдром, при котором после иисультообразного начала развивалось своеобразное расстройство мышечного тонуса на одной половине тела в виде упорной тонической судороги, распространяющейся на половину тела и в особенности резко выраженной в конечностях при отсутствии или слабой выраженности гемипареза и отсутствии пирамидных знаков. Автор назвал этот симптомокомплекс «апоплектической гемитонией» и объясняет его «болезненным гнездом на уровне мозговых узлов по соседству с задним отделом внутренней капсулы». Клиническое значение этого симптомоком-плекса подтверждено дальнейшими наблюдениями. Когда В. М. Бехтерев описал эту форму заболевания, еще не были известны клинические различия между поражениями пирамидного пучка и поражениями экстрапирамидной системы. В дальнейшем (1927) В. М. Бехтерев на основании анализа клинической симптоматики пришел к выводу, что при этой форме имеет место острое заболевание стриопаллидарной системы, вероятнее всего по типу очагового сосудистого процесса.
9. «О подчревном рефлексе» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 2).
В статье В. М. Бехтерев описывает новый рефлекс, на который до того авторы не обращали внимания. Ввиду того что уровень замыкания дуги этого рефлекса, по Бехтереву, расположен в наиболее низкой части грудного отдела спинного мозга, ниже чревного рефлекса, то исследование подчревного рефлекса, наряду с надчревным и чревным рефлексами, дает возможность точнее определять уровень поражения.
10. «О насильственном смехе в виде приступов, сопровождающихся тонической судорогой и чувством щекотания в левой руке» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, № 10).
459
В. М. Бехтерев приводит редкое и оригинальное клиническое наблюдение, которое объясняет патологическим процессом, локализующимся в области, соседней со зрительным бугром. Наблюдение представляет большой интерес.
П. «Об акромиальном или верхушечном рефлексе» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 3).
В. М. Бехтерев описывает в статье новый рефлекс с верхней конечности, который чаще всего наблюдается при повышенной рефлекторной возбудимости и должен быть причислен к периостальиым рефлексам.
12. «О чувствительности к постукиванию скуловой дужки как при знаке органического поражения основных частей мозга и его оболочек» (Обозрение психиатрии, неврологии и экспериментальной психологин, 1902, № 11).
В. М. Бехтерев установил, что глубокое постукивание через плессиметр скуловой дужки обнаруживает повышенную чувствительность к постукиванию, преимущественно на стороне болезненнвго процесса, в частности, при сифилитическом поражении основания мозга и его оболочек.
13. Pseudomelia paraesthetica как симптом черепно-мозгового поражения в области п. lenticularis (Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 12).
В. М. Бехтерев описывает своеобразный симптом, названный им pseudomelia paraesthetica и выражающийся в ложном ощущении движения парализованной конечности при полной неподвижности последней; коррекция зрения не всегда устраняет ложные ощущения. Этот симптом обусловлен поражением проводников мышечно-суставного чувства.
14. «Об особом сгибательном рефлексе ножных пальцев» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 13).
Статья касается описанного впервые В. М. Бехтеревым в 1901 г. рефлекса, наблюдающегося при органических поражениях пирамидных путей и получившего в литературе наименование Мендель-Бехтеревского рефлекса. Однако его следует по праву называть рефлексом Бехтерева, так как Курт Мендель описал тот же рефлекс на 3 года позже (в 1904 г.). Приоритет В. М. Бехтерева в описании этого рефлекса был признан самим Менделем, заявившим, что он не был знаком с работой В. М. Бехтерева.
15. «О насильственном плаче при мозговых поражениях» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 11).
В. М. Бехтерев был одним из первых, проанализировавших явления насильственного плача и смеха при мозговых поражениях (1885 г.). Им подчеркивается роль зрительного бугра в развитии этих симптомов.
16. «О рефлекторных явлениях при органических параличах центрального неврона и о новом тылыго-стопном сгибательном рефлексе стопы» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1911, № 7).
В. М. Бехтерев описывает новый рефлекс, выражающийся в тыльном сгибании стопы и пальцев (особенно большого) при повторном постукивании молоточком по тылу стопы. Этот рефлекс помогает отличать органические заболевания нервной системы от функциональных и является признаком поражения центральных двигательных путей. Автор касается здесь н других описанных им симптомов: клонуса стопы, тыльного раз-гибательного рефлекса стопы и пальцев, т. е. рефлекса, описанного значительно позднее также Мари и Фуа. В соответствии с этим тыльно-разги-бательный рефлекс стопы должен называться рефлексом Бехтерева.
17. «О рефлекторной эпилепсии под влиянием звуковых раздражений» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915, № 10—12).
Рефлекторная эпилепсия может быть вызвана под влиянием различных кожных, световых, звуковых раздражений. В. М. Бехтерев описы-
460
вает в статье редкий случай рефлекторной эпилепсии, в котором эпилептические приступы вызывались музыкой. Автор приводит подробное самонаблюдение больного.
18. «О двигательном и чувствительном феномене локтевого нерва» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1926, № 3).
В. М. Бехтерев описывает в этой статье двигательный и чувствительный феномен, наблюдаемый при механическом раздражении локтевого нерва и характеризующий собой повышенную возбудимость нервных стволов. Указанный феномен является симптомом смешанного характера, результатом как прямого механического раздражения нерва, так и рефлекторного эффекта. Он является более чувствительным, чем симптом Хвостека, и может зависеть, с. одной стороны, от гормональных влияний (спазмофилия), и с другой— от особенной иннервации двигательного церебрального пути.
19. «Тахифемия, тахибазия и пароксизмальное топтание на месте, как постэнцефалитический симптом» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1926, 24, № 1).
В. М. Бехтерев обратил внимание на маловыясненный постэнцефалитический синдром, «характеризующийся прогрессирующим или временным ускорением темпа движений». У одних больных расстройство выражалось во все ускоряющейся речи (тахифемия); у других также постепенно, но быстро ускорялась походка (тахибазия); у третьих, наконец, этот синдром выражался в непреодолимом многократном переступании с ноги на ногу (топтание на месте). В текущей литературе эти феномены были почти не освещены, а последний описывается В. М. Бехтеревым впервые. В. М. Бехтерев объясняет эти симптомы поражением «внепира-мидных двигательных проводников».
20. «О сложных рефлекторных явлениях в нижних конечностях при поражениях центрального двигательного неврона (рефлексы спинномозгового автоматизма)» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1927, № 1).
В этой статье В. М. Бехтерев приводит ряд описанных им рефлексов: разгибательный рефлекс ноги, сгибательный рефлекс ноги, приводящий рефлекс ноги. Автор делает анализ явлений суммирования н нарастания рефлексов. Приводятся убедительные доказательства приоритета В. М. Бехтерева в отношении рефлексов спинномозгового автоматизма и в том числе так называемых защитных рефлексов Мари и Фуа, описанных последними на 10 лет позже В. М. Бехтерева (Бехтерев описал сгибательный рефлекс ноги в 1900 г., а Мари и Фуа — в 1910 г.).
21. «Временное оцепенение или затормаживание активных движений как последствие поражения подкорковых мозговых узлов» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1927, № 2).
В статье В. М. Бехтерев впервые описал у больных с явлениями паркинсонизма своеобразный симптом, который выражается во внезапном временном застывании (затормаживании или оцепенении) членов или даже всего тела в определенном положении в периоды активных движений. Автор полагает, что здесь дело идет о временном возбуждении внепирамидных проводников.
IV. РАЗДЕЛ ПСИХИАТРИИ
1. «Случай полной слепоты, сопровождаемой галлюцинациями зрения» (Протоколы заседаний Общества психиатров в С.-Петербурге 10/Х 1883 г.).
Это короткое сообщение интересно тем, что показывает В. М. Бехтерева в качестве повседневного участника работы Общества психиатров в Петербурге. Обращает на себя внимание не только тщательность и яркость описания картины заболевания, но и применение сочетания неврологического и патофизиологического анализа. Следует отметить, что в период времени, к которому относится эта демонстрация В. М. Бехтерева, он работал над вопросом о значении органов равновесия в происхождении пространственных представлений н искажении последних. В опубликованных им работах по этому вопросу нет клинических иллюстраций. Приведенное описание показывает, что его теоретические положения основываются на тщательном изучении клинического материала, примером которого является описываемый редкий случай заболевания.
2. «О лечении навязчивых идей гипнотическим внушением» (Врач^ 1892, № 1).
В своей лечебной практике В. М. Бехтерев широко использовал к пропагандировал метод гипнотического внушения при различных нервных и психических заболеваниях. Данная статья является одной из первых среди многочисленных работ В. М. Бехтерева в этом направлении. Если достаточно популярной была идея лечения гипнозом истерии, то в отношении невроза навязчивых состояний в то время доминировала пассивная позиция, связанная с представлением о навязчивых состояниях как дегенеративных или конституциональных проявлениях.
Успешное применение гипноза при лечении ряда навязчивых состояний опрокидывало этот неправильный взгляд.
3. «О слышании собственных мыслей»(Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 10).
В. М. Бехтерев один из первых дает прекрасное описание клинической картины психического автоматизма в форме слышания собственных мыслей, т. е. в слухо-речевой сфере. Явления автоматизма встречались также в зрительной и двигательной сферах.
Первые наблюдения В. М. Бехтерева, на которые он ссылается в данной работе, относились к периоду времени незадолго до того, когда психические автоматизмы были описаны В. X. Кандинским (1890).
В. М. Бехтерев предвосхитил ряд наблюдений Клерамбо, в частности, эхо мыслей, мысли-подсказки и т. д., что дает основание считать более правильным обозначение синдрома психического автоматизма не синдромом Кандинского-Клерамбо, а синдромом Кандинского-Бехтерева.
В. М. Бехтерев приводит ряд ценных наблюдений и теоретические соображения, которые в настоящее время имеют актуальное значение. Он правильно относил слышание мыслей к слуховым галлюцинациям.
4. «О внушающем влиянии слуховых обманов чувств» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, №11).
В этой небольшой статье В. М. Бехтерев делает смелую и оригинальную попытку объяснить ту непреодолимую силу очевидности, какую имеют обманы чувств при слуховом галлюцинозе у больных с сохранным интеллектом и способностью критически разбираться во всем, что не соприкасается с содержанием галлюцинаторных переживаний.
Значение данной работы заключается в том, что устанавливаемая В. М. Бехтеревым связь некритического отношения больных к содержаниям галлюцинаций с состоянием, характерным для гипнотического внушения, дает ценный материал для понимания галлюцинаций в свете учения И. П. Павлова о гипнотических фазах.
5. «Об искусственном вызывании обманов чувств у алкоголиков, страдающих галлюцинаторными формами помешательства» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 2),
Из этой статьи видно, что В. М. Бехтерев один из первых начал заниматься изучением галлюцинаторных явлений, прибегая к дополнительным клиническим исследованиям. Он убедительно отвергает на основании собственных данных периферический генез этих явлений (не отрицая совсем известной роли периферического компонента) и содействует тем самым разработке этой проблемы. Он делает попытку подойти к ге-незу галлюцинаций с патофизиологической стороны. В. М. Бехтерев подчеркивает значение монотонных раздражений органов чувств для возникновения галлюцинаций.
6. «О периодических приступах ретроактивной амнезии» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 4).
Вопрос о сущности ретроактивной (ретроградной) амнезии принадлежит к числу наименее выясненных в психопатологии. Ретроактивная амнезия имеет разнообразные варианты, знание которых необходимо для решения более общих вопросов, связанных с этим явлением. Один из таких вариантов содержится в дайной работе В. М. Бехтерева.
Короткая ретроградная амнезия, возникающая приступообразно, с последующей амнезией на самый приступ представляет собой интереснейший вид периодической смены психопатологических явлений, не нашедший отражения в литературе как до описания В. М. Бехтеревым, так и после него.
7. «Об особом,развивающемся в детстве нервном поражении, выражающемся двигательными расстройствами и слабоумием» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, № 3).
В этой статье В. М. Бехтерев описывает редкую форму заболевания, выражающуюся в сочетании двигательных нарушений типа дрожания конечностей и мозжечковой атаксии, 'расстройства речи и явлений слабоумия. Отмечая у больной ряд 'эндокринно-вегетативных нарушений, В. М. Бехтерев полагает, что в основе всего синдрома лежит поражение, мозжечка и коры головного мозга. В. М. Бехтерев обращает внимание на то, что, помимо наследственных моментов, в развитии данного заболевания имел значение ряд других этиологических факторов, в частности, дефекты внутриутробного развития, асфиксия в родах и т. д. Автор в отличие от Криза не остался в плену у идеи о фатальной роли наследственности, господствовавшей в то время.
8. «О навязчивых кишечных и пузырных кризах и о значении при них лечения внушением» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, № 3).
Статья представляет одну из первых работ В. М. Бехтерева, в которой он описывает развитие навязчивых состояний, непосредственно связанных с нарушением деятельности внутренних органов.
Особый интерес описываемых в данной статье историй болезни заключается в указании, что возникновение заболевания происходило при условии предшествовавшего нарушения функции внутреннего органа.
Однако проблема кортико-висцеральной патологии была успешно разработана К. М. Быковым и его учениками.
463
9. «О расстройствах в сфере чувственного восприятия у душевнобольных» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 4).
Хотя статья написана в психологическом плане с сугубо субъективным толкованием психопатологических явлений, тем по менее она представляет определенную ценность. В ней приведена история болезни с ярким описанием явлений так называемой соматической деперсонализации. Ослабление чувственного восприятия у душевнобольных В. М. Бехтерев объясняет сноподобными психическими состояниями и отвлечением внимания. Этим дается ценный клинический материал к патофизиологическому пониманию указанного расстройства в свете созданного впоследствии Н. Е. Введенским и И. П. Павловым учения о фазовых явлениях, составляющих основу сноподобного состояния и открытого И. П. Павловым закона индукции нервных процессов, лежащего в основе внимания. Тот факт, что имя В. М. Бехтерева не упоминается в различных обзорах как в зарубежной, так и в отечественной литературе, посвященных проблеме деперсонализации и связи ее с расстройствами ощущений, является существенным пробелом и свидетельством изживаемой лишь сейчас недооценки выдающихся работ наших со-о течественников.
10. «О галлюцинаторном психозе, развивающемся при поражении органа слуха» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1S03, № 2).
Вопрос о роли поражения периферических органов (висцеральных органов чувств) при психических заболеваниях, обсуждавшийся старыми психиатрами, незаслуженно игнорировался в течение последних десятилетий. В настоящее время этот вопрос вновь привлекает интерес советских психиатров. Этому способствуют многочисленные работы отечественных физиологов и особенно К -М. Быкова и его сотрудников, показавшие теснейшую обоюдостороннюю зависимость между деятельностью внутренних органов и высшей нервной деятельностью. Разрабатывающаяся в настоящее время проблема рефлекторного патогенеза психических заболеваний при поражениях периферических органов, делает существенно важной клиническую казуистику, относящуюся к этой области.
Публикуемая работа В. М. Бехтерева содержит прекрасные клинические иллюстрации и обобщенную характеристику галлюцинаторных картин, сочетающихся с поражением органа слуха.
Замечание В. М. Бехтерева о значении личного (не наследственного) предрасположения, характеризующегося «особой возбудимостью нервной системы, благодаря чему периферическое раздражение со стороны органа слуха поднимает возбудимость центров до появления в них болезненной реакции, выражающейся развитием иллюзий и галлюцинаций», является прогрессивной точкой зрения отечественной науки начала XX века.
11. «О нарушении чувства времени у душевнобольных» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, N° 3).
В этой статье В. М. Бехтерев описывает у психически больных расстройство чувства времени, которое может наблюдаться вместе с другими психопатологическими симптомами или проявиться отдельно.
В. М. Бехтерев подчеркивает единство и взаимообусловленность элементарных и сложных психических нарушений.
12. «О значении взимания в отношении локализации и развития галлюцинаторных образов» (Обозрение психиатрии, неврологии и экспериментальной психологин, 1904, № 11).
Среди работ, которые В. М. Бехтерев посвятил вопросам клиники и патогенезу галлюцинаций, настоящая работа является одной из важнейших. Она посвящена изучению наиболее трудного вопроса — о проекции галлюцинаторных образов. Можно утверждать, что В. М. Бехтерев является первым автором, систематически исследовавшим этот вопрос. Ему удалось.обнаружить весьма важные закономерности. В современных
464
зарубежных трудах по вопросу о галлюцинациях (исключением в этом отношении является монография Мурга) эта работа В. М. Бехтерева не упоминается.
В настоящей статье В. М. Бехтерев показывает роль, которую играют внешние раздражители в проекции галлюцинаторных явлений. Задолго до Хэда и Холмса он отметил, что больные в некоторых случаях локализуют аффективное состояние в той или иной периферической области тела. В. М. Бехтерев рассматривает это явление как результат патологической связи между двумя различными сферами восприятия. Он пишет: «Есть основание полагать, что и патологические аффективные состояния, и общие ощущения могут при известных условиях относиться к тем или другим областям, где их локализация представляется явно ненормальною. Этим, например, объясняется заявление больных, что они испытывают «страх в мочевом пузыре» или что у них «тошнит левую ногу». При этом более внимательные расспросы показывают, что дело идет в этом случае не о фигуральном лишь выражении, а о таких состояниях, которые больные иначе выразить не могут. Он дает объяснение этому явлению в связи с общей своей концепцией об условиях проекции галлюцинаций.
13. «Признание действительности за галлюцинаторное переживание, как одно из проявлений психического расстройства» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1906, № 6).
В этой работе обнаруживается интерес В. М. Бехтерева к вопросам, относящимся к проблеме деперсонализации. Представленные им клинические примеры отличаются своеобразными проявлениями, а описания их— яркостью и интересной трактовкой.
В. М. Бехтерев не пользуется термином «деперсонализация», несмотря на то, что, судя по литературе, на которую он ссылается, проблема эта была ему хорошо известна.
В этой работе В. М. Бехтерев высказал свой взгляд на сущность соответствующих нарушений, который незаслуженно игнорировался последующими авторами.
14. «О галлюцинаторных воспоминаниях» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1907, № 6).
В работе содержится ряд интересных психопатологических примеров своеобразных галлюцинаторных переживаний и устанавливается связь их возникновения с различными нарушениями бодрственного состояния. В. М. Бехтерев, несомненно, прав, указывая, что описанные им галлюцинаторные явления проливают свет на патогенез галлюцинаций.
15. «Об отношениях, между психическими и нервными болезнями» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1908, № 1).
Статья посвящена обоснованию неразрывной связи, существующей между психическими и нервными болезнями. В. М. Бехтерев в данной работе прямо заявляет, что «психическая деятельность находится в полной и безусловной зависимости от деятельности высших корковых центров нервной системы...».
В. М. Бехтерев указывает на необходимость сближения психиатрии с остальной медициной. Он подчеркивает, что биохимические изменения в организме, ведущие к самоотравлению его, могут «вызывать функциональные расстройства деятельности мозговых клеток».
В статье содержатся отдельные устарелые положения. Так, В. М. Бехтерев не изменяет существовавшего тогда взгляда на эпилепсию как на иевроз. Истерию он трактует здесь еще как болезнь обмена.
Высказываемое В. М. Бехтеревым в данной статье положение, что в основе психических заболеваний лежат нарушения «функции высших центров нервной системы и ее сочетательных связей», близко подходит к учению И. П. Павлова о патофизиологии высшей нервной деятельности.
30 В. М, Бехтерев
465
Будучи выдающимся исследователем морфологии центральной нервной системы, В. М. Бехтерев хорошо понимал важнейшее патогенетическое значение функциональных нарушений в развитии психозов.
В. М. Бехтерев признает, что нужно «совершенно отказаться от анатомического принципа в объяснении разнообразных психических расстройств, наблюдаемых при душевных болезнях». В. М. Бехтерев подчеркивает, что «основа душевных или психических болезней должна быть патологофизиологическая, а не патологоанатомическая». Эти взгляды В. М. Бехтерева, противостоящие укоренившимся в медицине вирхови-анским представлениям о природе болезней, приобретают особое значение в настоящее время.
16. «О маниакально-меланхолическом психозе» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1909, № 11).
Значение статьи заключается в том, что в ней В. М. Бехтерев сумел преодолеть господствующие в психиатрической литературе, особенно за рубежом, взгляды Крепелина на маниакально-депрессивный психоз.
B. М. Бехтерев не признает решающей роли наследственности при этом заболевании. Он обнаруживает переходящие биохимические изменения в организме при маниакально-депрессивном психозе, как и при других психических болезрях, и тем самым ие оставляет места для моргано-вейс-манистских представлений об «эндогении». Полагая, что психоз вызывается самоотравлением организма, более выраженным при меланхолии, В. М. Бехтерев указывал на возможность его излечения. Утверждение В. М. Бехтерева, что некоторые воздействия на нервную систему приводят «к ее возбуждению, а затем к последовательному развитию угнетения и паралича», обнаруживает его стремление выяснить патофизиологическую основу маниакально-депрессивного психоза.
17. «О терапевтическом значении воспитания сочетательных рефлексов при истерических анестезиях и параличах» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1917, № 1—2).
В этой статье впервые ставится вопрос о применении методики воспитания условных (сочетательных) рефлексов как терапевтического метода при лечении истерических симптомов.
18. «О психорефлекториой терапии» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1914, № 10—11).
В. М. Бехтерев делает первые по времени попытки теоретического обоснования метода условнорефлекторной терапии нервных заболеваний. Им высказывается основное положение, что данный метод лечения должен содержать воздействия, направленные иа торможение образовавшихся патологических рефлексов и восстановление, возбуждение нормальных рефлексов. В связи с этим рассматривается и психотерапевтическое воздействие как словесный раздражитель, вызывающий необходимую для лечебных целей перестройку условнорефлекторной деятельности. В этой работе В. М. Бехтерев впервые выдвинул идею условнорефлекторной терапии алкоголизма. Последнее тем более заслуживает внимания, что американские ученые стремятся приписать себе приоритет условно-рефлекторной терапии алкоголизма.
19. «О сочетательно-рефлекторной терапии» (Вестник современной медицины, 1925, № 11).
Доклад, сделанный на съезде по охране здоровья детей и подростков, состоявшемся 16/XI 1925 г.
В. М. Бехтерев суммировал данные своих работ о сочетательнореф-лекторной терапии ряда болезненных состояний — анестезий, парезов и параличей при истерии, ночного недержания мочи, клептомании, алкоголизма.
В дальнейшем условнорефлекторная терапия находит приложение в работах Н. В. Канторовича, И. Ф. Случевского, И. В. Стрельчука,
C. П. Воробьева и др.
, Значение этих работ В. М. Бехтерева заключается не только в примененной им методике и описанных им формах, но и в учете самого услов-
466
норефлекториого момента в патогенезе заболевания и использовании его как одного из существенных принципов лечения.
20. «Pseudopolymelia paraesthetica в форме мнимых членов» (Обозрение психиатрии, неврологии и экспериментальной психологии, 1926, № 4—5)
В этой статье В. М. Бехтерев описывает новый симптом. Он справедливо сближает описанное им нарушение с галлюцинациями, монокулярной диплопией и полиопией, с кожной парестезией, явлениями фантома. Во всех нарушениях В. М. Бехтерев видит ложную пространственную проекцию ощущений вследствие недостаточности анализа нарушенных ощущений в мозговых центрах. Эта работа имеет большое значение. Эти идеи В. М. Бехтерева позже были развиты рядом авторов в учении о расстройстве схемы тела. Только в последнее время психиатрия становится на путь сближения расстройств схемы тела и галлюцинаций, без чего ни те, ни другие не могут быть поняты, на путь, начатый В. М. Бехтеревым как в этой работе, так и в ряде других его работ.
30*
систематический указатель
трудов и выступлений
академика в. м. бехтерева,
напечатанных
на русском языке1
СОСТАВЛЕН О. Б. КАЗАНСКОЙ и Т. Я. ХВИЛИВИЦКИМ
Примечание. В систематический указатель включено 613 наименований работ В. М. Бехтерева и 369 его выступлений, напечатанных на русском языке. Все работы В. М. Бехтерева, напечатанные на иностранных языках, как правило, за единичными исключениями, дублируют публикации в русских изданиях. В связи с этим работы на иностранных языках не включены в указатель.
ѓ. ОБЩАЯ БИОЛОГИЯ
Биологическое развитие мимики. Вестник психологии, 1911, в. 2, стр. 1—33.
Биологическое развитие мимики с объективно- психологической точки зрения.
Вестник знания, 1910, № 1, сто. 11—19; № 2, стр. 202—210; № 3, стр. 320—332; № 4, стр. 439— 446. То же, отд. изд., СПБ, 1910, 38 стр.
Биохимические системы и их роль в развитии организмов. Русский врач, 1 913, № 7, стр. 205— 210. То же, отд. изд. СПБ, 1913, 24 стр.
Нормальная анатомия и гистология
Книги
Проводящие пути мозга.
Ученые записки Казанского университета, 1892, ноябрь—декабрь, стр. 55—122, Библиография, стр. 176—190. То же отд. изд. Казань, 1893, 192 стр.
Проводящие пути мозга. Казань, 1893, 196 стр.
Проводящие пути спиииого и головного мозга.
Руководство к изучению внутренних связей мозга, СПБ, 1896— 1898, ч. 1—2, часть 1. 390 стр., ч. 2, 383 стр. То же, ч. 1,
М. — Л., 1926, 496 стр.
Статьи
Заметка по поводу работ Л. О. Дарк-шевича о проводнике светового раздражителя в сетчатой оболочке глаза на глазодвигательный иерв.
Значение гормоиизма и социального отбора в эволюции организмов. Природа, 1916, № 10, стр. ИЗО— 1158.
Предисловие к кн.: Н. А. Белова, Физиология типов. Под редакцией В. М. Бехтерева. Орел, 1924, 247 стр.
Роль социального отбора в эволюции видов.
Вестник знания, 1926, № 13, стр. 841—844.
Социальный отбор и его биологическое значение.
Вестник знания, 1912, № 12, стр. 947—955.
Архив психиатрии, неврологии и судебной психопатологии, 1889, т. 12, № 1, стр. 1—13.
К вопросу о вторичных перерождениях мозговой ножки. Вестиик клинической и судебной психиатрии и невропатологии, 1885, в. 1, стр. 210—224. Реф.: Архив психиатрии, неврологии и судебной психопатологии. 1885, т. 6, № 1, стр. 143.
К вопросу о мозговых полосках продолговатого мозга (striae medul-lares acusticae).
Медицинское обозрение, 1892,т. 37, № 5, стр. 470—478.
К вопросу о наружных сочетательных волокнах мозговой коры. Медицинское обозрение, 1891, т. 35, № 22, стр. 926—928.
К вопросу о составных частях задних столбов спинного мозга. Медицинское обозрение, 1887,т. 28, № 17, стр. 427—429.
К вопросу о центральных окончаниях слухового иерва и о физио-
II. АНАТОМИЯ И ФИЗИОЛОГИЯ
471
логическом значении его преддвер-ной ветви.
Вестник клинической и судебной психиатрии и невропатологии, 1887, в. 1, стр. 217—222.
Мозг Д. И. Менделеева.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1908, № 10, стр. 637—639. Совместно с Р. Л. Вейнбергом.
Мозговые полушария.
В кн.: Основания к изучению микроскопической анатомии человека и животных. Под ред. М. Д. Лавдовского и Ф. Овсянникова, т. 2, СПБ, 1888, стр. 905—933 с библиографией.
Направления зрительных волокон на пути от коленчатых тел до четверохолмия.
Вестник клинической и судебной психиатрии и невропатологии, 1883, в. 1, стр. 128—130.
Несколько слов по вопросу об отношении сосудов межпозвоночных узлов к содержащимся в них нервным клеткам. Неврологический вестник, 1896, т. 4, в. 2, стр. 141 — 142.
Новый случай перерождения волокон наружной части основания мозговой ножки (так называемого пучка Turck'a).
Русская медицина, 1885, № 33, стр. 609—611.
Реф.: Архив психиатрии, неврологии и экспериментальной психологии, 1885, т. 6, № 3, стр. 56.
О вторичном перерождении нисходящего пучка передней иожки мозжечка центрального пучка покрышки и петлевого слоя. Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 8, стр. 602—603.
О выделении различных систем спинного мозга по методу развития. Неврологический вестник, 1901, т. 9, в. 2, стр. 169—191.
О двойном лучепреломлении нервными волокнами.
Вестник клинической и судебной психиатрии и невропатологии, 1889, в. 2, стр. 155.
О двух пучках, входящих в состав внутреннего отдела задней ножки мозжечка, и о развитии волокон слухового иерва.
Врач, 1885, № 25, стр. 408—409. Реф.: Протоколы заседаний Обще-
472
стваврачей-психиатров за 1885 г., СПБ, 1886, стр. 125.
О двух пучках, составляющих среднюю ножку мозжечка. Врач, 1885, № 9, стр. 131 — 132.
О задних корешках, месте их окончания в сером веществе спииного мозга и об их центральном продолжении.
Вестник клинической и судебной психиатрии и невропатологии, 1887, в. 1, стр. 203—214.
О малоизвестной системе волокон, проходящих по периферии передне-наружного отдела шейной части спиниого мозга.
Неврологический вестник, 1900, т. 8, в. 4, стр. 65—68.
О направлении суживающих зрачков волокон в головном мозгу и о локализации центра для радужной оболочки и для сокращений глазных мышц.
Вестник клинической и судебной психиатрии и невропатологии, 1883, в. 1, стр. 89—114. Реф.: Труды заседаний Общества русских врачей за 1882—1883 гг., стр. 113.
Дополнение к статье «О направлении суживающих зрачок волокон в головном мозгу». Протоколы заседаний Общества психиатров в С.-Петербурге за 1883 г., М., 1885, стр. 57.
О неполном перекрещивании зрительных нервов в chiasma высших млекопитающих. Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 10, стр. 744.
О нервных путях спинного и головного мозга.
В кн.: Основания к изучению микроскопической анатомии человека и животных. Под ред. М. Д. Лавдовского и Ф. Овсянникова, т. 2, СПБ, 1888, стр. 940—961.
О нисходящих связях зрительного бугра.
Неврологический вестник, 1904, т. 12, в. 2, стр. 209—216.
О переднем внутреннем пучке боковых столбов спииного мозга. Неврологический вестник, 1900, .т. 8, в. 3, стр. 152—155.
О петлевом слое иа основании результатов исследования по способу развития.
Неврологический вестник, 1895, т. 3, в. 3, стр. 103—120.
О продольных волокнах сетевидной фармации на основании исследования их развития и о соединениях сетчатого ядра покрышки. Врач, 1886, № 6, стр.. 112.
О развитии клеток мозговой коры у человека.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 8, стр. 609—611.
О соотношении нервных клеток при посредстве деидритов. Отчеты научных собраний врачей Q.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 47.
О составе задних столбов спинного мозга на основании истории их развития.
Врач. 1884, № 51, стр. 855—856. Реф : Архив психиатрии, неврологии и судебной психопатологии,
1885, т. 5, № 2—3, стр. 267.
О составных частях веревчатого тела (corpus restiforme). Вестник клинической и судебной психиатрии и невропатологии,
1886, в. 1, стр. 25—33.
Реф.: Архив психиатрии, неврологии и судебной психопатологии,
1887, т. 8, № 1—2, стр. 208—209. О составных частях передней ножки
мозжечка.
Вестник клинической и судебной
психиатрии и невропатологии,
1886, в. 2, стр. 217—225.
О составных частях так называемой остаточной области боковых столбов спинного мозга. Врач, 1885, № 29, стр. 473—474.
О центральных окончаниях блуждающего нерва и о составе так называемого одиночного пучка продолговатого мозга.
Вестник клинической и судебной психиатрии и невропатологии,
1887, в. 2, стр. 13—21.
О центральных окончаниях тройничного нерва.
Вестьик клинической и судебной психиатрии и невропатологии, 1887, в. 1, стр. 215—216.
Об изменениях спинного мозга вслед за перерезкой нервных корешков. Протоколы заседаний общества психиатров за 1884 г., СПБ, 1885, стр. 3—6.
Совместно с П. Я. Розеибахом..
Об «оливчатом» пучке шейной части спинного мозга.
Неврологический вестиик, 1894, т. 2, в. 2, стр. 117—122.
Об особенном пучке длинных волокон, проходящем в передней части боковых столбов спинного мозг?.. Протоколы заседаний Общества психиатров за 1884 г., СПБ, 1885, стр. 48—49.
Об особой промежуточной системе волокон, заложенных в боковых пирамидальных пучках. Неврологический вестник, 1895, т. 3, в. 3, стр. 93—96.
Об особенной составной части боковых столбов спиниого мозга и о начале волокон большого восходящего корешка тройничного нерва. Врач, 1885, № 26, стр. 422—423.
Об особенном, до сих пор не описанном, пучке, проходящем в верхнем этаже стволовой части мозга. Протоколы заседаний Общества психиатров в СПБ, за 1885 г., СПБ, 1886, стр. 12—13.
Об особом внутреннем пучке боковых столбов.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 4, стр. 265—267
Об особом ядре сетевидного образования на уровне отделов варо-лиева моста.
Неврологический вестник, 1898, т. 6, в. 2, стр. 182—183.
Об относительном развитии и различных положениях пирамидальных пучков у человека и животных и о содержании в этих пучках волокон, отличающихся более ранним развитием.
Медицинское обозрение, 1890, т. 34, № 13—14, стр. 108—112.
Об ядрах глазодвигательных нервов (п. oculomotorii, abducentis и tro-chlearis) и о связи их друг с другом. Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 7, стр. 515—522.
Связи мозговой коры с nucleus lenticularis.
Обозрение психиатрии, неврологии и экспериментальной психоли-гии, 1900, № 11, с. 860—86L Реф.: Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900—1901 гг., стр. 7—8.
47'i
Экспериментальные исследования относительно перекреста волокон зрительного нерва. Еженедельная клиническая газета, 1883, № 2, стр. 22—25; № 3, стр. 38—41.
Реф.: Протоколы заседаний Общества психиатров в С.-Петербурге за 1882 г. СПБ, 1883, стр. 15.
Выступления в прениях по поводу сообщений
Базилевский А. П., О свежих вторичных перерождениях после перерезки задней мозжечковой иожки и поражения мозжечка. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 32.
Бергер В. М., О состоянии слюноотделительного аппарата у новорожденных животных. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 52.
Б и р ю л я-Б е л ы н и ц к и й Ф. А. Несколько случаев перерыва центральной борозды. Протоколы заседаний Общества психиатров за 1906—1907 гг., СПБ, 1907, стр. 52—53.
Б л умен ау Л. В., Препараты спинного мозга по Гольджи. Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 4—7.
Вырубов Н. А., Резорцин как заместитель осмиевой кислоты при обработке мозга по Гольджи, Протоколы заседаний Общества психиатров за 1897 г., СПБ, 1899, стр. 37—38.
Вырубов Н. А., К вопросу о центральных окончаниях и связях VII и VIII пары черепных нервов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и иервиых болезней за 1899—■ 1900 гг., СПБ, 1900, стр. 42—43.
Г е р в е р А. В., Об окончаниях в коре мозга латерального пучка основания мозговой иожки. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 23.
Г е р в е р А. В., О центральных началах отводящего нерва. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899—1900 гг., СПБ, 1900, стр. 19—20.
Гизе Э. А., О нервных клетках спинного мозга человека (по способу Гольджи).
Отчеты научных собраний врачей клиники душевных и нервных болезней за 1897—1898 гг., СПБ, 1899, стр. 40—41.
Гизе Э. А., О развитии невроглии в спинном мозгу человека. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 87—88.
Гизе Э. А., Об овальном пучке в поясничном утолщении спинного мозга.
Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 107.
Добротворский М. С, Демонстрация вторичных перерождений в спинном мозгу при перерезках в грудной области. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 30.
Жуковский М. Н., К учению о тонком строении нервной клетки и ее отростков.
Отчеты научных собраний врачей .С.-Петербургской клиники душевных и нервных болезней за 1904—1905 гг., СПБ, 1905, стр. 12.
Жуковский М. Н., Об анатомических связях лобиых долей. Отчеты научных собраний врачей С.-Петербургской клиники нервных душевных и болезней за 1895—1896 и 1896—1897 гг., СПБ, 1898, стр. 70.
Кауфман П. Ю., О центростремительных нервах артерий. Отчеты научных собраний врачей СПБ клиники душевных и нервных болезней за 1907—1908 гг., СПБ, 1909, стр. 37.
Корольков П. Я-, Верхний (в варолиевом мосту) перекрест двигательных (пирамидных) путей и отношение их к ядрам черепных нервов у некоторых грызунов.
474
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 9.
Корольков П. Я-, Верхний (в варолиевом мосту) перекрест двигательных (пирамидных) путей и отношение их к ядрам черепных нервов у человека. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908-— 1909 и 1909—1910 гг., СПБ, 1910, стр. 31.
Корольков П. Я-, О клетках центральной первой системы и симпатической нервной системы, окрашенной по способу Ehrich'a и их взаимном отношении. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 31.
Корольков П. Я-, Об окончании разветвлений в печени и в слюнных железах. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—• 1896 и 1896—1897 гг., СПБ, 1898, стр. 93—94.
Лазурский А. Ф., О петлевом слое.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ; 1898, стр. 74—75.
Осипов В. П., О центральных окончаниях IX пары иервов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 85—86.
Осипов В. П., «О центральных окончаниях п. Vagi. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 42.
П а с с е к В. П., Новый способ окрашивания нервных клеток. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 8.
П и л ь ц, Центральные пучки двигательных иервов глаза. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 32—33.
П у с с е п Л. М., Демонстрация препаратов мозжечка. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 67.
Рахманов А. В., К вопросу об окраске неврофибрилл. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1904— 1905 гг., СПБ, 1905, стр. 50—51.
Рахманов А. В., Неврофибрил-лии и хромотофильное вещество в нервных клетках.
Отчеты научных собраний врачей С.-Петербургской клиники нервных и душевных болезней за 1905—1906 и 1906—1907 гг., СПБ, 1908, стр. 27—28.
Реймерс Н. К., О перерождении в спинном мозгу вслед за перерезкой задних и передних ко решков.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 103—104.
Р у б а ш к и н В. Я., О строении невроглии.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902— 1903 гг., СПБ, 1903, стр. 46—47.
Свенторжецкий Б. А., Случай аномалии роландовой борозды.
Протоколы заседания Общества психиатров за 1894 г., СПБ, 1896, стр. 9.
С и н а н и Б. Н., О направлении центростремительных волокон рефлекторной дуги, суживающих зрачок.
Протоколы заседаний Общества психиатров за 1883 г., СПБ, 1885, стр. 47—49.
С н е с а р е в П. Е., К вопросу о волокнах Rhinencephalon'a у лягушки.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905—
475
1906 и 1906—1907 гг., СПБ, 1908, стр. 71—72.
С н е с а р е в П. Е., О маргинальной глии.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 80.
Телятник Ф. К-, О перекресте зрительных нервов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 61—62.
Телятник Ф. К.,0 перерождениях, наступающих в центральной нервной системе после разрушения верхнего червячка мозжечка. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 76—77.
Телятник Ф. К., О технике способа окраски центральной нервной системы по Marchi. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 59.
Ткаче н ко М. П., Проводящие пути мозжечка по методу развития.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902— 1903 гг.; СПБ, 1903, стр. 16—17.
Т р о ш и н Г. Я-, К вопросу о связях островка.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902— 1903 гг., СПБ, 1903, стр. 118.
Т р о ш и и Г. Я-, О поясном пучке (cingulum).
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902— 1903 гг., СПБ, 1903, стр. 59—60.
Т р о ш и н Г. Я-, Центральные связи чувствительных черепных нервов у человека. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 87.
Шипов Н. Н., Демонстрация макротома.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 56.
Шипов Н. Н., О связях свода. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 59—60.
Эрнст Э. Э., О нисходящих связях зрительного бугра со средним мозгом и мозговым стволом. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 48.
Ю р м а н Н. А., Анатомические и физиологические исследования черного вещества.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 21—22.
Патологическая анатомия и гистология
Статьи
Демонстрация двух мозгов — одного с обширными разрушениями мозговой поверхности, обусловленными атрофическим склерозом и другого с бугоркоьой опухолью на дне IV желудочка.
■ Протоколы заседаний Общества.
психиатров в С.-Петербурге за 1895— 1896 гг., СПБ, 1897, стр. 53—54.
Демонстрация мозга больного, страдавшего апраксией. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 26—30.
Демонстрация мозга больной, страдавшей мозговой опухолью. Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 3, стр. 220—221.
Демонстрация мозга больного, страдавшего парасимболией. Протоколы заседаний Общества психиатров за 1,912 г., СПБ, 1912, стр. 13—14.
Демонстрация мозга больного с парасимболией.
Отчеты научных r-обраний врачей С.-Петербургской клиники душев-
476
ных и нервных болезней за 1910— 1911 гг., СПБ, 1911, стр. 20—21. Демонстрация мозга больного, умершего от новообразования в области верхней части продолговатого мозга и нижнего отдела варо-лиева моста.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 45—46. Демонстрация случая сирингомиэ-лии.
Протоколы заседаний Общества психиатров в С.-Петербурге за 1895—1896 гг., СПБ, 1897, стр. 44. Новые наблюдения одеревенелости позвоночника с патологоанатоми-ческим исследованием. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 5, стр. 392—403. О перерождении волокон в продолговатом мозгу при нарастающем параличном слабоумии (dementia paralytica).
Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 12, стр. 928—929. О порозе мозга.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней, 1900— 1901 гг., СПБ, 1901, стр. 59—62. О результатах микроскопического исследования сифилитических поражений мозговой ткани. Протоколы заседаний Общества психиатров в С.-Петербурге за 1880 г.,СПБ, 1881, стр. 6—15. О сифилитическом поражении мозговой ткани.
В ки.: Новейшие исследования по различным отраслям медицинских наук, СПБ, 1880, стр. 139—186. Поражение корешков caudae equinae вследствие прорастания нижней части позвоночного столба злокачественным новообразованием. Неврологический вестник, 1893, т. 1, в. 3, стр. 50—70. Предъявление мозга с разрушением передних и внутренних частей обеих височных долей. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 7, стр. 589—590. Результаты патологоаиатомического вскрытия трупа больного с одеревенелостью позвоночника.
Отчеты научных собраний всачей С.-Петербургской клиники душевных и нервных болезней за 1897—-1898 гг., СПБ, 1899, стр. 23—24. Случай местной атрофии мозговой поверхности, обусловившей судорожные припадки с отведением глаз и головы на одну сторону. Еженедельная клиническая газета, 1881, № 1, стр. 8—12; № 2, стр. 24—29; № 3, стр. 37—47.
Выступления в прениях по поводу сообщений
Бабкин, Влияние искусственных швов черепа у молодых животных на его рост и развитие. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 39—40. Б л у м е н а у Л. В., Микроскопические изменения в мозгу одной душевнобольной.
Протоколы заседаний Общества психиатров за 1893 г., СПБ, 1895, стр. 32„33, 34.
Вендерович Е. Л., Демонстрация способа изучения иа непрерывной серии срезов свежих иевроносистемиых изменений в головном мозгу человека. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1910—— 1911 гг., СПБ, 1911, стр. 24.
Вырубов Н. А., Демонстрация препарата множественной эмболии мозга.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 33.
Вырубов Н. А., К патологической анатомии множественного неврита.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 23—24.
Вырубов Н. А., Случай опухоли основания мозга. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899,, стр. 9.
Г е р в е р А. В., Патологоанатоми-ческие изменения мозговой коры при остром безумии.
477
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 37.
Гуревич М. О., О резистентности фибриллярного аппарата нервных клеток при патологических условиях.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907— 1908 гг., СПБ, 1909, стр. 22—23.
Данилло С. Н., Демонстрация патологоанатомических препаратов центральной нервной системы. Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ,
1897, стр. 54—55, 61, 62. Жуков Н. А., О влиянии голодания новорожденных на развитие центральной нервной системы. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ,
1898, стр. 13.
Жуков М. Н., Демонстрация мочга микроцефала. Протоколы заседаний Общества психиатров за 1902—1903 гг., СПБ, 1904, стр. 34.
Жуков М. Н., К вопросу о патологической анатомии острого бреда.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 21.
Жуковский М. Н.,. К патологической гистологии амиотро-фического бокового склероза с бульварным началом. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 40.
Жуковский М. Н., О восходящих и нисходящих вторичных перерождениях при компрессионном миэлите нижней шейной и верхней грудной областей. Отчеты научных собраний врачей С.-Петербургской клиники душевных и иервиых болезней за 1901 — 1902 гг., СПБ, 1902, стр. 85.
Жуковский М. Н., Предъявление микроскопических препаратов
sclerosis disseminata.
Отчеты научных собраний врачей С.-Петербургской клиники душев-
ных и нервных болгзней за 19С1 — 1902 гг., СПБ, 1902, стр. 3.
Корольков П. Я., Опухоль мозга в области четверохолмия у 11-месячного ребенка. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 91.
Корольков П. Я-> Склероз сосудов коры мозга в связи с идиотизмом.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 56.
Лазурский А. Ф., О патологоанатомических изменениях мозговой коры при старческом ослаблении.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 31—32.
Любимов С. Я-. К вопросу о патологических изменениях мозга при status epilepticus. Протоколы заседаний Общества психиатров за 1899 г., СПБ, 1901, стр. 54.
М а щ е н к о С. М., Патологоана-томические изменения в мозговой коре при вторичном слабоумии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 57.
Мержеевский И. П., К патологии идиотизма. Протоколы заседаний Общества психиатров за 1900—1901 гг., СПБ, 1902, стр. 15.
Никитин М. П., Демонстрация препаратов молочной железы. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ, 1908,. стр. 15—16.
Н и л ь с о н Е. А., Патологоанато-мические изменения нервной системы при хроническом отравлении этиловым алкоголем животных.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ,. 1908, стр. 31—32.
478
Осипов В. П., Демонстрации аномалии головного мозга. Протоколы заседаний Общества психиатров за 1895 — 1896 гг., СПБ, 1897, стр. 101.
Осипов В. П., Патологоанато-мические изменения нервной системы, вызываемые поясничным проколом.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 6—7.
П а с с е к В. П., Патологоанатоми-ческие изменения в центральной нервной системе под влиянием лучей радия.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 91—92.
П е к е р В. А., Об изменениях в мозгу в зависимости от искусственной ишемии. Протоколы заседаний Общества психиатров за 1884 г., СПБ, 1885. стр. 18.
Петров Е. П., Об изменениях нервных клеточек при остром отравлении алкоголем и сивушным маслом.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 43.
П у с с е п Л. М., О влиянии перевязки и прижатия брюшной аорты иа изменения в спинном мозгу. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 70.
Рахманов А. В., Патологоана-томические исследования случая прогрессивной мышечной атрофии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 111 — 112.
Рыбалкин Я. В., Случай кровоизлияния варолиева моста с частичным параличом чувствительности.
Протоколы заседаний Общества психиатров за 1893 г., СПБ, 1895, стр. 27—28. Т р о ш и н Г. Я-, Случай опухоли четверохолмия.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 43—44.
Толочинов И. Ф., Об изменениях ядер четверохолмия моста и продолговатого мозга при прогрессивном параличе помешанных. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 82—83.
Фальк М. А., Микроскопические изменения почек и печени в 52 случаях психоневрозов. Протоколы заседаний Общества психиатров за '1895—1896 гг., СПБ, 1897, стр. 98.
Фальк М. А., Об изменениях почек и печени при прогрессивном параличе и слабоумии на почве органического поражения мозга. Протоколы заседаний Общества психиатров за 1894 г., СПБ, 1896, стр. 25—26.
Физиология
Книги
Мозг и его деятельность. М. — Л., 1928, 327 стр.
К физиологии равновесия тела (отправление центрального серого вещества третьего мозгового желудочка).
Воеино-медицинский журнал, 1883* кн. 7, стр. 178—231. То же, отд. изд., СПБ, 1883, 55 стр.
О локализации сознательной деятельности у животных и человека,. СПБ, 1896, 62 стр.
О связях так называемых периферических органов равновесия с мозжечком.
Русская медицина, 1884, № 3,. стр. 61—64; № 4, стр. 85—86; № 5, стр. 110—113. То же, отд. изд., СПБ, 1884, 30 стр.
Основы учения о функциях мозга* в. 1—7, СПБ, 1903—1907.
Физиология двигательной области мозговой коры.
Архив психиатрии, неврологии и судебной психопатологии, 1887, т. 9, № 2, стр. 57—67; № 3, стр. 25—41; т. 10, № 1, стр. 1—64. То же, отд. изд., Харьков, 1887, 90 стр.
47θ
Статьи
Воздействие на поведение животных путем непосредственного (бессловесного) внушения. Вопросы изучения и воспитания личности, 1921, № 3, стр. 471—478.
Заднее двухолмие как центр слуха, голоса и отраженных движений. Неврологический вестник, 1895, т. 3, в. 2, стр. 63—74. Реф.: Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 11, стр. 857—858.
Значение сочетания методов развития и перерождения с живосечениями для экспериментальной физиологии нервной системы и о роли нежных и мозжечковых пучков спинного мозга в отношении функций равновесия. Неврологический вестник, 1895, т. 3, в. 1, стр. 97—104.
Значение эпителиальных приборов, чувствующих нервов в отношении качественного различия воспринимаемых ощущений. Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 2, стр. 89—93.
Изменение функциональной деятельности нервной системы в зависимости от действия кожных раздражителей.
В сб. Новое в рефлексологии и физиологии нервной системы, в. 1, 1925, стр. 254—290. Совместно с Г. Е. Шумковым.
Изучение функций предлобных и других областей мозговой коры с помощью сочетательно-двигательных рефлексов. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915; № 4, 5, 6, стр. 336—347; № 7, 8, 9, стр. 353— 356.
Исследование вынужденных или насильственных движений у животных при разрушении стенок воронкообразной части третьего желудочка в мозгу. Еженедельная клиническая газета, 1882, № 7, стр. 97—102; № 8, стр. 119—124; № 11, стр. 161 — 165.
Исследования над зрительным центром мозговой коры. Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 1, стр. 71—72.
Исследование функций мозговой ко ры с помощью естественных соче тательных рефлексов. Значение этого метода по отношению к центрам внутренних органов и различных секреций. Обозрение психиатрии, неврологии и экспериментальной психологии, 1908, № 8, стр. 471—479.
К вопросу о влиянии мозговой коры и зрительных бугров на глотание. Неврологический вестник, 1894, т. 2, в. 2, стр. 130—137.
К вопросу о соотношении друг с другом клеток центральной нервной системы.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 3, стр. 200.
К вопросу о функции мозжечка. Медицинское обозрение, 1890, т.34, № 13—14, стр. 102—108.
К вопросу об иннервации желудка. Медицинское обозрение, 1890, т.33, № 2, стр. 185—189. Совместно с Н. А. Миславским. Реф. (Н. И. Ковалевский). Архив психиатрии, неврологии и судебной психопатологии, 1890, т. 15, № 8, стр. 123.
К физиологии межпозвоночных узлов. Об изменениях спииного мозга вслед за перерождением нервных корешков.
Вестник клинической и судебной психиатрии и невропатологии, 1884, в. 1, стр. 155—170. То же, Врач, 1884, № 9, стр. 129, реф. Протоколы заседаний Общества психиатров за 1884 г., СПБ, 1884, стр. 3—8. Совместно с П. Я. Ро-зенбахом.
О влиянии гипноза и внушения в нем на простой и сочетательные рефлексы.
Труды Государственного института медицинских знаний, в. 1, Л., 1927, стр. 181—227. Совместно с В. Н. Мясищевьгм.
О влиянии голодания иа новорожденных животных, в особенности на вес и развитие их мозга. Неврологический вестник, 1895, т. 3, в. 1, стр. 85—96, реф. (Грейденберг Б.). Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 10, стр. 775—776.
480
О влиянии коры большого мозга на акт глотания и на дыхание. Неврологический вестник, 1894, т. 2, в. 2, стр. 123—129. Совместно с П. А. Останковым.
О влиянии мозговой коры на отделение слюны. .
.Медицинское обозрение, 1888, т. 30, Л1' 13, стр. 52—54. Совместно с Н. Л. Миславским. Реф. (П. И. Ковалевский). Архив психиатрии, неврологии н судебной психопатологии, 1888, т. 12, ЛЬ 3, стр. 115.
О влиянии мозговой коры человека на сердцебиение, давление крови и дыхание.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № И, стр. 854—857.
О влиянии мозговой коры и центральных областей мозга па давление крови и деятельности сердца. Архив психиатрии, неврологии и судебной психопатологии, 1886, т. 8, № 3, стр. 1—5. Совместно с Н. А. Миславским.
CJ влиянии травматического воспаления коры головного мозга на ее возбудимость.
Неврологический вестник, 1894, т. 2, в. 4, стр. 171 — 173.
О влиянии удаления мозговых полушарий на зрение и слух. Протоколы заседаний Общества психиатров в С.-Петербурге за 1883 г., М., 1885, стр. 30—32.
О возбудимости двигательных цен-
■ тров мозговой коры у новорожденных щенят. Врач, 1886, № 34, стр. 609—610.
Реф. 1. (Рахманов). Медицинское обозрение, 1887, т. 27, „\l> 1, стр. 80—81.
2. Архив психиатрии и неврологии судебной психопатологии, 1887, т. 8, ЛЬ 1—2, стр. 209.
О / возбудимости мозговой коры у новорожденных животных. Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, ЛЬ 7, стр. 487—489.
О возбудимости различных частей головного мозга у новорожденных животных. Врач, 1889, ЛЬ 15, стр. 353—356.
О вынужденных движениях, обнаруживающихся при разрушении мозговой коры-
Русская медицина, 1885, ЛЬ 1, стр. 6—8, № 3, стр. 54—55.
О вынужденных или насильственных движениях у животных при разрушении некоторых частей центральной нервной системы. Труды Общества русских врачей в С.-Петербурге с, приложением протоколов заседаний общества за 1881 — 1882 гг., СПБ, 1882, стр. 161 — 168.
О двигательных корковых центрах обезьяны.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 88.
То же. Ученые записки Казанского университета, 1900, кн. 1, стр. 18—19.
О закономерности в воспитании и развитии двигательных сочетательных рефлексов.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1916, ЛЬ 5—6, стр. 36—45.
О значении исследования местных рефлексов при изучении функций мозговой коры.
Неврологический вестник, 1908, т. 15, в. 3, стр. 443—458.
О зрительной области мозговой коры и об отношении ее к глазным мышцам.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, ЛЬ 4, стр. 257—273, ЛЬ 5, стр. 336—356.
О зрительной площади на поверхности мозговых полушарий. Архив психиатрии, неврологии и судебной психопатологии, 1890, т. 15, ЛЬ 1, стр. 1—32.
О корковом зрительном центре. Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, № 8, стр. 375—379.
О корковых центрах сужения и расширения зрачков в задних частях полушарий обезьян. Обозрение психиатрии, неврологии п экспериментальной психологии, 1899, ЛЬ 7, стр. 555—558.
О /юкализации зрачкового центра и о направлении суживающих зрачок волокон в головном мозгу. Труды Общества русских врачей в С.-Петербурге за 1882—1883 гг., СПБ, 1883, стр. 113—126.
31 Ь. М. Бсхтепсг.
181
О локализации зрительного центра коры на внутренней поверхности затылочной доли у собак. Обозрение психиатрии, неврологии и экспериментальной психологии, 1911, № 8, стр. 449—453.
О локализации кожных (осязательных и болевых) ощущений и мышечного сознания на поверхности мозговых полушарий. Врач, 1883, № 30, стр. 465—467.
О мозговых центрах движения влагалища у животных. Медицинское обозрение, 1891, т. 36, № 15, стр. 247—259. Совместно с Н. А. Миславским.
О направлении суживающих зрачок волокон в головном мозгу и о локализации центра для радужной оболочки и для сокращения глазных мышц.
Протоколы заседаний Общества психиатров в 1882 г., СПБ, 1883, стр. 15.
О последовательной смене явлений возбуждения и торможения при воспитании сочетательных двигательных рефлексов. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914 — 1915, Кя 10, 11, 12, стр. 549—554.
О последовательной смене явлений возбуждения и торможения при воспитании сочетательных двигательных рефлексов. Обозрение психиатрии, неврологии и экспериментальной психологии, 1916, № 5, 6, стр. 46—52.
О произвольном расширении зрачка. Неврологический вестник, 1895, т. 3, в. 1, стр. 23—43.
О расположении двигательных центров мозговой коры у человека по результатам раздражения их фарадическим током при мозговых операциях.
Неврологический вестник, 1896, т. 7, в. 3, стр. 1—6.
О результатах исследования возбудимости заднего отдела лобной доли. Неврологический вестник, 1899, т. 7, в. 1, стр. 172—175.
О слуховых центрах в коре полушарий.
Неврологический вестник, 1899, т. 7, в. 3, стр. 138—154.
О соединении верхних олив и об их вероятной физиологической роли. Врач, 1885, № 32, стр. 526—528.
Реф. Архив психиатрии, невроло-гии и судебной психопатологии, 1885, т. 6, № 3, стр. 54—56.
О сочетательно-рефлекторных изменениях мозгового пульса. Труды Государственного института медицинских знаний, в. 4, Л., 1928, стр. 3—29. Совместно с В. Н. Мясищевым.
О способах воспитания цепи сочетательных рефлексов. В кн.: Вопросы изучения и воспитания личности, год 4-й, 1927, № 3—4, стр. 3—9. Совместно с Н. И. Добротворской.
О суживающих зрачок и аккомода-тивных центрах мозговой коры. Неврологический вестник, 1900, т. 8, в. 1, стр. 207—209.
О так называемом судорожном центре и о центре передвижения тела на уровне варолиева моста. Неврологический вестник, 1896, т. 4, в. 4, стр. 97—101.
О центрах дивергенции и конвергенции в головном мозгу. Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 7, стр. 482—488.
О центрах мочевого пузыря. Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 8, стр. 586—589.
О центральной и периферической иннервации кишок. Труды Общества естествоиспытателей при Казанском университете, 1889, т. 20, стр. 215—278. Совместно с Н. А. Миславским. »
О чувствительной п двигательной роли зрительного бугра. Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 2, стр. 92—100.
О чувствующей функции так называемой двигательной области мозговой коры у человека. Неврологический вестник, 1899, т. 7, в. 3, стр. 6—10.
О явлениях, наблюдаемых при разрушении различных частей! нервной системы у новорожденных животных и о развитии у них мозговых функций. Медицинское обозрение, 1890, т. 33, № 4, стр. 392—397. Реф. (Н. И. Мухин). Архив в психиатрии, неврологии и судебной психопатологии, 1890, т. 15, N° 3, стр. 125.
482
О явлениях, обнаруживающихся у животных вслед за перерезкою, задних пучков спинного мозга и об отношении последних к функциям равновесия. Вестник клинической и судебной психиатрии и невропатологии, 1889, в. 1, стр. 107—125. Реф. (Н. И. Ковалевский). Архив психиатрии, неврологии ft судебной психопатологии, 1889, т. 15, № 1, стр. 129.
О явлениях, следующих за перерезкою зрительных волокон виу-тримозговых полушарий (в соседстве заднего отдела внутренней капсулы).
Вестник клинической и судебной психиатрии и невропатологии, 1883, в. 2, стр. 206—215.
Об измерении объема мозга.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 10, 11, стр. 660— 661.
Об иннервации и мозговых центрах слезоотделения.
Медицинское обозрение, 1891, т. 35, № 12, стр. 1170—1175. Совместно с Н. А. Миславским.
Об исследовании возбудимости отдельных пучков в спинном мозгу новорожденных животных. Врач, 1887, N° 22, стр. 435—437.
Об отправлении зрительных бугров. Врач, 1883, № 4, стр. 51—53, № 5, стр. 68—70.
Реф. Протоколы заседаний Общества психиатров в С.-Петербурге за 1883 г., СПБ, 1883, стр. 3—7.
Об отправлении зрительных бугров у животных и человека. Вестник клинической и судебной психиатрии и невропатологии, 1885, в. 2, стр. 91 — 173. Реф. Архив психиатрии, неврологии н судебной психопатологии, 1886, т. 8, № 1, стр. 112.
Об отправлении олиечзтых тел продолговатого мозга. Врач, 1882, № 34, стр. 566—567; № 35, стр. 586—588.
Об отправлении четверохолмия. Врач, 1883, № 32, стр. 497—499; № 33, стр. 515—517; N° 34, стр. 532—534; № 35, стр. 547—548.
Реф. Протоколы заседаний Общества психиатоов С.-Петербурга за 1883 г.," СПБ, 1884, стр. 27—30.
Опыты над перерезкой слухового нерва и полукружных каналов уха.
Труды Общества русских врачей в СПБ, 1882—1883 гг., СПБ, 1882, стр. 9—26.
Развитие полового влечения с точки зрения рефлексологии. Русский врач, 1918, № 29—32, стр. 169—175; N° 33—36, стр. 193— 199.
Результаты исследования произвольно вызываемых изменений сердечной деятельности. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896 — 1897 гг., СПБ, 1898, стр. 36.
Результаты опытов с раздражением задних отделов полушарий и лобной доли у обезьян. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг. СПБ, 1899, стр. 14—15.
Результаты перерезки слухового нерва.
Протоколы заседаний Общества психиатров в 1882 г., СПБ, 1883, стр. 12—14.
Своеобразное распределение паралича чувствительности и движения при поражении боковых частей нижнего отдела продолговатого мозга и переходной области между продолговатым и спинным мозгом. Неврологический вестник, 1894, т. 2, в. 1, стр. 1—42.
Теория соприкосновения (contact theorie) и учение о разрядах нервной энергии как условие проведения нервного возбуждения. Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 1, стр. 23—28.
Теория спинномозгового сосудисто-приспособительного рефлекса для объяснения трофизма наружных покровов.
Современная психоневрология,
Киев, 1925, N° 1, май, стр. 5—13.
Учение о нервных единицах (neuron) и соотношениях их друг с другом. Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 1, стр. 12—22.
Центр движения мочевого пузыря в головном мозгу.
31*
483
Архив психиатрии неврологии, и судебной психопатологии, 1888, т. 12, № 2, стр. 74—79. Совместно с Н. А. Миславским. Экспериментальные данные относительно мозжечка. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898—
1899 гг., СПБ, 1899, стр. 57—58. Экспериментальные данные относительно функций мозжечка. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 6, стр. 508—509.
Экспериментальные исследования над круговыми движениями у животных. Еженедельная клиническая газета, 1881, №34, стр. 585— 586.
Экспериментальные исследования об отправлении зрительных бугров. Протоколы заседаний Общества психиатров в С.-Петербурге в 1884 г., СПБ, 1885, стр. 3—7.
Экспериментальные исследования относительно перекреста зрительных волокон в chiasma ηη. opticorum. Протоколы заседаний Общества психиатров в 1882 г., СПБ, 1883, стр. 15.
Выступления в прениях по поводу сообщений
Агаджанянц К- С, О корковом центре зрения. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 50—54.
Акопенко А. Ф., Влияние ти-реоидэктомии на рост костей у молодых животных. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 83.
А с п и с о с Н. и А с п и с о в И., К вопросу о локализации корковых центров и о центральных проводниках верхней ветви его. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899—
1900 гг., СПБ, 1900, стр. 34—35. Афанасьев Н. И. К вопросу
о выработке сочетателыю-двига-7'ельных рефлексов.
Отчеты научных собраний врачей (".-Петербургской клиники душевных и нервных болезней за 1910— 1911 гг., СПБ, 1911, стр. 51.
Бари А. Э., О влиянии головного мозга на слюноотделение. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 71—72.
Бари А. Э., О возбудимости двигательной области коры мозга новорожденных животных. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 22—23.
Бе й нар К- А., Об изменениях мозгового кровообращения под влиянием острого алкогольного отравления.
Протоколы заседаний Общества психиатров за 1897 г., СПБ, 1899, стр. 08.
Б е л и ц к и й 10. К-, Корковые центры аккомодации. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 37—38.
Бонды рева О. И., Сочетательно-двигательный рефлекс на обонятельные раздражения у человека.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1909— 1910 гг., СПБ, 1910, стр. 95.
Боришпольский Е. С., Об изменениях мозгового кровообращения при вращении животных на горизонтальном круге. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 34.
Боровиков И. В., Случай расстройства равновесия при страдании полукружных каналов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1904— 1905 гг., СПБ, 1905, стр. 38—39.
В а л ь к е р К. И., Сочетательная двигательная реакция у собаки на цветные раздражения. Отчеты научных собраний врачей С -Петербургской клиники душевных и нервных болезней за 1908 —
484
1909 и ШОУ—1910 гг., СПБ, 1910, стр. 79.
Васильев, Кокаинизация спинного мозга как анестезирующее средство.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 57—58.
Введенский Н. Е., Возбуждение, торможение и наркоз. Протоколы заседаний Общества психиатров за 1900 и 1901 гг., СПБ, 1902, стр. 35—36.
Введенский Н. Е., К учению о центральной иннервации. Протоколы заседаний Общества психиатров за 1909—1910 гг., СПБ, 1911, стр. 10—11.
Введенский Н. Е., Об электрических свойствах нервов. Протокол заседания Общества психиатров за 1900—1901 гг., СПБ, стр. 50—51.
Г е й м а и Н. М., Рефлексы со слизистой оболочки полости рта на слюнные железы. Труды Общества русских врачей в СПБ, 1903, стр. 401—404.
Г е н к и и И. Л., Кожные гальванические токи как метод исследования некоторых функций мозга. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907— 1908 гг., СПБ, 1909, стр. 69.
Гервер А. В., О мозговых центрах ассоциированных движений глазных яблок.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 7—8.
Гервер А. В., К учению о тормозящих функциях мозговой коры. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 30—31.
Гервер А. В., О влиянии головного мозга на отделение желудочного сока.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 26.
Г и з е Э. А. и Л а з у р с к и й А. Ф., Дыхание и пульс в гипнозе. Отчеты научных собраний врачей С.-Петербургской клиники душев-
ных и нервных болезней sa 1899—
1900 гг., СПБ, 1900, стр. 6—7.
Г о л а н т Р. Я., Демонстрация электромиограмм при отравлении дигаленом и вератрином. Отчеты научных собраний врачей С.-Петербургской клиники душевных'н нервных болезней за 1910—1911 гг., СПБ, 1911, стр. 51-52.
Г о л а н т Р. Я., К выработке сочетательно-двигательного рефлекса на вкусовые раздражения. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 60—61.
Г о л а н т Р. Я-, О действиях си-нусообразных колебаний тока на двигательный нерв. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 58—59.
Горшков Я. П., О локализации центров обоняния в мозговой коре. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900—
1901 гг., СПБ, 1901, стр. 80—81. Горшков Я. П., О месте центра
вкуса в мозговой коре. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 45—46.
Г р е к е р Р. А., Двигательные явления, наблюдаемые при раздражении верхнего червячка мозжечка.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907— 1908 гг., СПБ, 1909, стр. 30—32.
Г р е к е р Р. А., Демонстрация собак с удаленными центрами отделения желудочного сока. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907— 1908 гг., СПБ, 1909, стр. 59—61.
Грибоедов А. С, О корковых потоотделительных центрах. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 —
1902 гг., СПБ, 1902, стр. 91—92.
-185
Д а м а с к и н Н. И., Механическая теория мышечной деятельности.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 36.
Жуков Н. А., О влиянии быстрой остановки мозгового кровообращения перерезкою аорты на возбудимость двигательных центров мозговой коры.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 10.
Жуковский М. Н., О влиянии мозговой коры и подкорковых узлов на дыхание. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 103.
II з р а э л ь с о н Ж. И., К вопросу о сочетательно-двигательном рефлексе на тактильное раздражение. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908-— 1909 и 1909—1910 гг., СПБ, 1910, стр. 56—57.
Израэльсон Ж. И., О вторичном сочетательно-двигательном рефлексе у собаки и человека. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, стр. 96—97.
Израэльсон Ж. И., О сочетательно-двигательном рефлексе на тепловое раздражение у собаки. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 н 1909—1910 гг., СПБ, стр. 37.
Израэльсон Ж. И., Топографическая локализация сочетательно-двигательного рефлекса на тактильное раздражение у собаки. Особенности сочетательно-двигательного рефлекса на составное раздражение (тактильное и тепловое) у человека.
•Отчеты научных собраний врачей С.-Петербургской клиники душевных н нервных болезней за 1908—
486
1909 и 1909—1910 гг., СПБ,
1910, стр. 83. Карпинский А. И., О влиянии коры головного мозга на почечное отделение.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1904— 1905 гг., СПБ, 1905, стр. 14—15.
Карпинский Л. И., О корковых центрах мочеотделения. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 51—52.
Краинский Н. В., О физической природе внешних раздражений органов чувств. Протоколы заседаний Общества психиатров за 1909—1910 гг., СПБ,
1911, стр. 4—5.
Л азу реки и А. Ф., О влиянии мышечных движений на мозговое кровообращение.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 83.
Ларионов В. Е., О корковых слуховых центрах у собак. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ,
1898, стр. 81—82. Левченко Г. В., О влиянии
снотворных средств на мозговое
кровообращение.
Протоколы заседаний Общества
психиатров за 1897 г., СПБ,
1899, стр. 03. Левченко Г. В., О методах
исследования кровообращения в мозгу.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 19—20.
Л е о и т о в и ч А. В. и Ларионов В. Е., Учение о нейронах. Труды II съезда отечественных психиатров в 1905 г., Киев, 1907, стр. 298—299.
Н а р б у т В. М., Влияние экстирпации мозгового придатка на мозговое кровообращение и обмен веществ.
Протоколы заседаний Общества психиатров за 1902 и 1903 гг. СПБ, 1904, стр. 76.
ii г. p бут В. М., Материалы к разработке гистологической теории сна.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898—
1899 гг., СПБ, 1899, стр. 74.
Н а р б у т В. М., Об электровозбудимости нервов и мышц у новорожденных.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 58—59.
11 а р б у т В. М., Состояние дендрита в коре головного мозга во время естественного сна. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899—
1900 гг., СПБ, 1900, стр. 38—39. Никитин М. П., О влиянии
головного мозга на отделение молока.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1904— 1905 гг., СПБ, 1905, стр. 48—49.
П и к и т и п М. П., О психомоторных центрах. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905—1906 гг. и 1906— 1907 гг., СПБ, 1908, стр. 3—4.
Ноишевский К. И., О вибрационном чувстве Treitel'H и костном чувстве Egger'a. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900—
1901 гг., СПБ, 1901, стр. 26—27. Ноишевский К. И., О железистой чувствительности. Протоколы заседаний Общества психиатров за 1900 и 1901 гг., СПБ, 1902, стр. 60—62.
Орлеанский Л. М., О состоянии мозгового кровообращения, вызванного механическою задержкою дыхания.
Отчеты научных собраний врачей (".-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 29.
Осипов В. П. и Ноишевский К- П., Волосковая чувствительность у здоровых. Отчеты научных собраний врачей С.-Петербургской клиники душев-
ных и нервных болезней за 1897 — 1898 гг., СПБ, 1899, стр. 44—46.
Осипов В. П. и Ноишевский К. П., О волосковой чувствительности у больных. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 49.
Павлов И. П., О р б е л и Л. Л. и Эльясов М. И., Прения по докладам, сделанным ими на Сеченовском заседании. Труды русских врачей п С.-Петербурге, 1908, т. 75, стр. 310—313.
Пассе к В. П., Нервные клетки спинного мозга в состоянии покоя и под влиянием электрического раздражения двигательных центров коры.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 60—61.
Певницкий А. А., Случаи навязчивых состояний, леченных по психоаналитическому методу. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907—
1908 гг., СПБ, 1909, стр. 40—41. Плохинский Н. М., О влиянии коры мозга и подкорковых узлов на сокращение матки у кролика.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 79—80.
Протопопов В. П., Влияние адреналина на черепно-мозювое кровообращение и на возбудимость психомоторных центров мозговой коры.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903 — 1904 гг., СПБ, 1904, стр. 43.
Протопопов В. П., Демонстрация собак с утратой местных кожных рефлексов после удаления чувство-двигательных центров мозговой коры.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—
1909 и 1909—1910 гг., СПБ., 1910, стр. 28.
487
Протопопов В. П., К вопросу об анатомическом пути сочетательных рефлексов.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 134—135.
Протопопов В. П., Об условиях образования специфического сочетательного рефлекса на звук, об анатомической дуге сочетательных двигательных рефлексов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 51—53.
Пуссен Л. М., Влияние прижатия брюшной аорты на мозговое кровообращение.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898—
1899 гг., СПБ, 1899, стр. 42. II у с с е п Л. М., Влияние совокупления на мозговое кровообращение у самок.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899—
1900 гг., СПБ, 1900 г., стр. 73—74. Пуссеп Л. М., О влиянии рентгеновских лучей на возбудимость мозговой коры.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 63.
Пуссеп Л. М., О корковых центрах эрекции. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 40—41.
С. о к о в и ч М. К-, О влиянии tuber cinereum на температуру тел а.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 57—58.
Соболевский А. В., Демонстрация нового прибора для исследования температурной чувствительности.
Отчеты научных собраний врачей С.-Петербургской клиники душев-
ных и нервных болезнен за 190? — 1908 гг., СПБ, 1909, стр. 24—25.
Соболевский А. В., О влиянии кровопускания на мозговое кровообращение.
Отчеты научных собраний врачей С.-Петербургской клиники дун.ев-ных и нервных болезней за 1899 -1900 гг., СПБ, 1900, стр. 82.
Сол ух И. П., О проницаемости кожных покровов для света вольтовой дуги.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899--1900 гг., СПБ, 1900, стр. 51-52.
С и и р то в И. М., О влиянии музыки на кровяное давление. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ, 1908, стр. 8—9.
С п и р т о в И. Н., Демонстрация коркового центра слюноотделения. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907 — 1908 гг., СПБ, 1909, стр. 52- 58.
Срезневский В. В., Об изменении скрытого периода коленного рефлекса под влиянием исп\ га. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905—1906 и 1906—1907 гг., СПБ, 1908, стр. 20—21.
С т р а х о в и ч И. В., О токах высокого напряжения и большой частоты. Их физические свойства вообще и по отношению к организму животных и человека. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 01.
Тарханов И. Р., Об иннерйа-ции светящегося аппарата (к физиологии сна).
Протоколы заседаний Общества психиатров за 1900 и 1901 гг., СПБ, 1902, стр. 42—43.
Телятник Ф. К. и Б о р и ш-польский Е. С, О влиянии чувственных раздражений на черепно-мозговое кровообращение. Отчеты научных собраний врачей С.-Петербургской клиники душенных и нервных болезней за 1897— 1898 гг , СПБ, 1899, стр. 12—13.
488
Телятник Ф. К., О возбудимости нервной системы при уремии.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895-— 1896 и 1896—1897 гг., СПБ, 1898, стр. 55—56.
Телятник Ф. К., Теоретические и практические соображения по поводу исследования мозгового кровообращения посредством измерения кровяного давления в центральном и периферическом конце сонной артерии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897—
1898 гг., СПБ, 1899, стр. 6. Торопов Н. К-> Зрительная
реакция собак при удалении затылочных долей больших полушарий. Протоколы заседаний Общества русских врачей в С.-Петербурге, 1908, т. 75, март—май, стр. 454.
Трапезников А. В., О центрах глотания.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 72.
Т р и в у с С. А., О влиянии цветового освещения иа пулос. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 44—45.
Т р и в у с С. А., Токи действия в коре под влиянием периферических раздражений. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898—
1899 гг., СПБ, 1899, стр. 73. Френкель Р. А., К вопросу
о щекотном чувстве. Сочетательно-двигательная реакция иа щекотание.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—
1909 и 1909—1910 гг., СПБ, 1910, стр. 91.
Ч а г о в е ц В. Ю., Демонстрация опытов с искусственным сном \ животных под влиянием прерывистого гальванического тока Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ. 1908, стр. 14—15.
Ч а г о в е ц В. Ю., О приложении новых электрохимических теории к электрофизиологии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ. 1898, стр. 66.
Чудновский Я- Б., К вопрос\ о путях проведения волосковон чувствительности. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 74—75.
Ш а й к е в и ч М. О., Физиологические исследования чечевичной ядра.
Протоколы заседаний Обществ;, психиатров за 1902 и 1903 гг.. СПБ, 1904, стр. 66—67.
Ш т и д а В. В., О так называсмы\ тепловых центрах в головном мозгу. Протокол заседания Обществ;: психиатров за 1902 и 1903 гг.. СПБ, 1904, стр. 55.
Щеглов А. Л., Изменения черепно-мозгового кровообращения под влиянием антипирина и пирамидона.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болгзней за 1901 — 1902 гг., СПБ, 1902, стр. 21—22.
Ющенко А. И., О желудочное пищеварении, преимущественно of отделительной работе желудочных желез у душевнобольных. Протоколы заседаний Общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 19.
III. НЕВРОПАТОЛОГИЯ И ПСИХИАТРИЯ
Общие вопросы
Книги
Невропатологические и психиатрические наблюдения, СПБ, 190С, 299 стр.
Невропатологические и психиатрические наблюдения, СПБ, 1910. 114 стр.
Объективное исследование нервно-психической деятельности. Речь, произнесенная 4 сентября 1907 г.
48t
на Международном конгрессе по психиатрии, неврологии, психологии и общественному призрению душевнобольных, СПБ, 1908, 26 стр.
Статьи
Витковский о неврологии.
Вестник клинической и судебной психиатрии и невропатологии, 1883, в. 2, стр. 258—259.
Война и психозы. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915 гг., № 4, 5, 6, стр. 317—335.
Война и психозы.
В кн.: Вопросы мировой войны Пг., 1915, стр. 590—604.
Вопросы вырождения и борьба с ним. Обозрение психиатрии, неврологии и экспериментальной психологии, 1908, № 9, стр. 518—521.
Международный конгресс по психиатрии, неврологии, психологии и общественному призрению душевнобольных, происходивший в Амстердаме от 2—7 сентября 1907 г. Обозрение психиатрии, неврологии и экспериментальной психологин, 1908, № 4, стр. 251—253.
О возможности применения рентгеновских лучей к нервным и душевным болезням.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 гг. и 1896—1897 гг., СПБ, 1898, стр. 37—38.
О насильственном смехе в виде приступов, сопровождающихся тонической судорогой и чувством щекотания в левой руке. Обозрение психиатрии, неврологии и экспериментальной психологии, 1901,- № 10, стр. 721—725.
О применении сочетательно-двигательных рефлексов как объективных приемов исследования в клинике нервных и душевных болезней.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1910, № 8, стр. 449—471.
О расторжении брака по сумасшествию. .
Обозрение психиатрии, неврологии и экспериментальной психологин, 1906, № 3, стр. 161 — 169. Совместно с М. С. Добротворским.
Об отношении между психическими и нервными болезнями. Обозрение психиатрии, неврологии и экспериментальной психологии 1908, № !, стр. 12—22. Реф. И. Жилин, Неврологический вестник, 1908, т. 15, в. 2, стр. 410.
Основные задачи психиатрии, как объективной науки. Русский ьрач, 1912, № 6, стр.181 — 189.
Предисловие к кн.: В. Маньян, Легрен, Поль Морис. Вырождающиеся (исследование). Перевод с франц. Ю. В. Португалова, СПБ, 1903, 163 стр.
Предисловие к кн.: А. Форель, Половой вопрос. Перевод с нем. Р. М. Марковича, т. 1—2, СПБ, 1911.
Применение метода сочетательно-двигательного рефлекса к исследованию притворства. Русский врач, 1912, № 14, стр. 461.
Психо-биологические вопросы, Научное обозрение, 1902, № 1, стр. 1—9; № 2, стр. 1 — 16; № 3, стр. 2—21; № 4, стр. 24; № 5, стр. 1—25; № 6, стр. 1—34; № 8, стр. 1—21.
Реф. (Ю. Португалов), Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 2, стр. 139—144.
Рецензии на работы.
Проф. И. Оршанского: Etude sur l'heredite normal ct morbide, st. Pet., 1894. Deux tipes de la fa-mille, Moscou, 1892.
Законы наследственности, Харьков, 1898.
Роль наследственности в передаче болезней, СПБ, 1897.
Наследственность и законы происхождения полов.
В кн.: Отчет о восьмом присуждении Академии наук премий митрополита Макария в 1899 г., СПБ, 1902, стр. 31—37.
Выступления по поводу сообщений
А р и н ш т е й н Л. С, Введение токсинов и антитоксинов в цере-брально-спинальную жидкость как метод экспериментальных исследований в невропатологии. Отчеты научных собраний врачей С.-Петербургской клиники душев-
490
ных и нервных болезней за 1903— •1904 гг., СПБ, 1904, стр. 35—36.
Ь с к, Демонстрация случая с болезнью костной системы. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903—-!904 гг., СПБ, 1904, стр. 13—14.
Бори ш польский Е. С,О состоянии кровообращения в головном мозгу во время припадков падучей.
Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 104—106.
В и х р е в Г., Прибор для стереоскопического рассматривания рентгеновских снимков. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 84.
Воротынский Б. И., О необходимости включения судебной психопатологии в систему юридического образования. Труды II съезда отечественных психиаторов в 1905 г., Киев, 1907, стр. 114 — 118.
Выступления в прениях по поводу доклада Психиатрической комиссии Общества невропатологов и психиатров, состоящего при Московском университете, о классификации душевных болезней для отчетов психиатрических учреждений и однообразной форме отчетов для этих учреждений. Труды II съезда отечественных психиатров в 1905 г., Киев, 1907, стр. 72—73, 80, 96.
Г о л а н т Р. Я. и Срезневский В. В., Демонстрация электрокардиограмм.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 97.
Гольцингер Ф. Ф., Впечатление врача-психиатра из путешествия по восточной экваториальной Африке.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 77—78.
Г о л ь ц и н г с р Ф. Ф., Кессонная болезнь. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899 — 1900 гг., СПБ, 1900 г., стр. 25,
Гольцингер Ф. Ф., Нервные болезни в Абиссинии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1897—1898 гг., СПБ, 1898, стр. 79.
Гольцингер Ф. Ф., Случай leprae maculoanaestheticae с демонстрацией больной. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 3.
Грибоедов А. С, Об искусственном вызывании падучего припадка у эпилептиков. Протоколы заседаний Общества психиатров за 1906—1907 гг., СПБ, 1907, стр. 66.
Дрознес М. Я., Важнейшие задачи современной практической психиатрии.
Труды II съезда отечественных психиатров в 1905 г., Киев, 1907, стр. 214.
Жуков Н. А., К вопросу о травматической эпилепсии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 9.
Жуков Н. А., О влиянии травматического воспаления мозговой коры на скрытый период возбуждения двигательных центров ее. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 8.
Иванов И. И., О физиологическом и патологическом аффекте в судебномедицинском отношении. Труды II съезда отечественных психиатров в 1905 г., Киев, 1907, стр. 197—199.
Карпинский А. И., О процессе самоотравления при миотомии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 66.
491
Корольков II. Я., Демонстрация больной с мик:едемою. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 55—56.
Корольков П. Я., Демонстрация ребенка, пораженного слизистым отеком (myxoedema). Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1S04, стр. 90—91.
Маляревский Е. X., Об исключении из школ детей с эпилептическими припадками. Протоколы заседаний Общества психиатров за 1900 и 1901 гг., СПБ, 1902, стр. 71.
Муратов В. А., Вопросы дня в постановке преподавания нервных и душевных болезней. Труды II съезда отечественных психиатров в 1905 г., Киев, 1907, стр. 230—231.
Н ю р е н б е р г А. Д., Экспериментальные данные о внутренней секреции hypophysis и их клиническое значение.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1910— 1911 гг., СПБ, 1911, стр. 37—38.
Ольдерогге В. В., Психозы в искусстве.
Протоколы заседаний Общества психиатров за 1898 г., СПБ, 1899, стр. 22.
Осипов В. П., Корковое происхождение падучих приступов, вызванных отравлением собак essence d'absinthe. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 13—14.
11 у с с е п Л. М., Демонстрация девятилетнего ребенка с микседе-мой.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 1.
Розенбах П. Я., Вопрос о расторжении брака вследствие помешательства одного из супругов. Протоколы заседаний Общества психиатров за 1898 г., СПБ, 1899, стр. 34.
Сербский В. П., О судсбпопси-хиатрической экспертизе. Труды II съезда отечественных психиатров в 1905 г., Киев, 1907, стр. 148.
Методы исследования и общая симптоматика нервных бслезней
Статьи
Временное оцепенение или затормаживание активных движений, как последствие поражения подкорковых мозговых узлов. Обозрение психиатрии, неврологии и рефлексологии, 1927, Л1> 2, стр. 113—118.
Гемитония и гемиклония как следствие гнездного мозгового поражения.
Неврологический вестник, 1897, № 3, стр. 13.
Реф. научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895 — 1896 и 1896—1897 гг., СПБ, 1898, стр. 49—51.
Гипертрофическое развитие грудных желез у мужчин, как приобретенное изменение.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1906, № 4, стр. 245—250. Двусторонний, периодически обостряющийся паралич главных мышц со своеобразными колебаниями в управлении движением верхних век.
Неврологический вестник, 1898, т. 6, в. 2, стр. 169—181.
Демонстрация больного с поражением задней половины спинного мозга.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1908, № 12, стр. 752—753.
Демонстрация больного с ревматическим поражением позвоночника. Обозрение психиатрии, неврологии и экспериментальной психологии,
1907, № 8, стр. 510—511. Демонстрация больного с tabes dor-
salis, страдающего периодической икотой.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902 стр. 72—74.
492
Демонстрации больного с трапскор-тикалыюй парафазией или пари-симболией.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907— 1908 гг., СПБ, 1909, стр. 61.
Демонстрация больного с черепным ранением.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1904—
1905 гг., СПБ, 1905, стр. 43—44. Демонстрация больной, страдающей
своеобразным расстройством в двигательной сфере кишечника. Протоколы заседаний Общества психиатров в С.-Петербурге за 1909—1910 гг., СПБ, 1911, стр.86— 88. Демонстрация мозга (два больных с корсаковским психозом: Первый — смерть от рака пищевода, второй — от туберкулеза легких). Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905—
1906 и 1906—1907 гг., СПБ, 1908, стр. 11 — 13.
Демонстрация мозга больного, страдавшего апраксией. Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 1, стр. 68.
Демонстрация мозга больного, умершего в Казанской губернской земской больнице и вскрытого в день заседания (алкоголик). Неврологический вестник, 1894, Лз 2, стр. 64 (приложение)
Демонстрация случаев с резко выраженным нарастанием рефлексов и необычными клоническими феноменами, с указанием на некоторые из недавно описанных рефлексов.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 2, стр. 49.
Демонстрация случая афазического расстройства.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1905, № 3, стр. 228—229. Демонстрация случая с нарастанием
рефлексов.
Обозрение психиатрии, неврологии
и экспериментальной психологии,
1906, № 1, стр. 63.
Демонстрация случая судороги в плечевом поясе в виде маятнико образного движения то одного, то другого плеча.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899—
1900 гг., СПБ, 1900, стр. 65—66. Демонстрация случая транскортикальной парафазии.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 86—89.
Демонстрация тылыюсгибательного рефлекса стопы.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1910— 1911 гг., СПБ, 1911, стр. 32—34.
Еще несколько слов об объективных признаках гиперестезии и анестезии при травматических неврозах. Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 3, стр. 161—162.
Исследования над развитием иадуч-ных приступов.
Неврологический вестник, 1894, т. 2, в. 3, стр. 173—178.
К вопросу о значении рентгенизации при огнестрельных ранениях головы.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 1, стр. 58.
К симптоматологии невропатической «одервенелости» позвоночника.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1916, № 5—6, стр. 3—6.
Наблюдения трофических изменений кожи в зависимости от расстройств нервной системы. Еженедельная клиническая газета, 1881, № 27, стр. 477—483, № 28, стр. 489—497.
Насильственный смех при отсутствии паралича личных нервов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900—
1901 гг., СПБ, 1901, стр. 9—10. Несколько слов по поводу «Заметки
об исследовании и значении так называемых сухожильных рефлексов», приват-доцент С. Н. Да-нилло. Врач, 1891, № 25, стр. 604—605.
493
Несколько случаев повреждения коры большого мозга. Медицинский вестник, 1879, № 27, стр. 217—218; №29, стр. 233—234; № 30, стр. 241—243; № 33, стр. 265—266; № 34, стр. 273—275; № 35, стр. 281—283; № 36, стр. 289—291; № 44, стр. 353— 355; № 45, стр. 361—363; № 46, стр. 371—373; № 47, стр. 373— 379; № 48, стр. 388—389; № 49, стр. 396—397; № 50, стр. 403—404; № 51, стр. 410.
Неудержимый или насильственный смех и плач при мозговых параличах.
Неврологический вестник, 1893, т. 1, в. 3, стр. 1—32.
Новый алгезимер или болемер. Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 6, стр. 408—413.
О безболезненном акроэритрозе. Обозрение психиатрии, неврологии и экспериментальной психологии, 1908, № 10, стр. 605—612.
О волосковой гиперестезии или три-хогиперестезии.
Обозрение психиатрии, неврологии и рефлексологии, 1900, № 8, стр. 572—584.
О выдыхательном рефлексе и об общем сгибательном рефлексе с тыла стопы.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 114 — 115.
О вызывании боли при седалищной невралгии гиперэкстензией конрч-ности.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1907, № 4, стр. 210—212.
О вынужденных движениях, обнаруживающихся при разрушении мозговой коры.
Русская медицина, 1885, № 1, стр. 6—8; № 3, стр. 54—55.
О глазном рефлексе.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, Кя 11, стр. 801—804.
О двигательном и чувствительном феномене локтевого нерва. Обозрение психиатрии, неврологии п рефлексологии, 1926, № 3, стр. 163—169.
О двух редких формах личного тика Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 12, стр. 895—897.
О запястно-фаланговом рефлексе. Обозрение психиатрии, неврологии и экспериментальной психологии,
1902, № 7, стр. 487—488.
О звуковой проводимости при постукивании костей черепа и позвоночника как метода исследования в нервных болезнях. Неврологический вестник, 1894, т. 2, в. 1, стр. 91 — 100.
О значении исследования механической возбудимости мышц и так называемых «мышечных» рефлексов в нервных болезнях. Обозрение психиатрии, неврологии и экспериментальной психологии,
1905, № 5, стр. 329—332.
О значении лопаточно-плечевого рефлекса.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 10, стр. 726—729.
О компенсаторных или заменяющих движениях при мозговых поражениях.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1903, № 10, стр. 733—737.
О лечении желудочных кризов спин-носухоточных.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1896, № 1, стр. 28—29.
О локализации двигательной апрак-сии.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1906, № 1, стр. 9—18.
О малоизвестных отраженных явлениях при нервных болезнях и о распознавательном значении так называемого стопного феномена и изменений со стороны сухожильных и кожных рефлексов. Неврологический вестник, 1895, т. 3, в. 2, стр. 75—88.
О малоизвестных ранних симптомах спинной сухотки. Обозрение психиатрии, неврологии и экспериментальной психологии,
1897, Л1> 9, стр. 650—652. О местной апраксии.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 12, стр. 942.
494
'Го же. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902—1903 гг., СПБ, 1903, стр. 50.
О мышечном валике при миотомии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 11, стр. 814—815.
О надглазничной нспрогрессирую-щей атрофии.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902— 1903 гг., СПБ, 1903, стр. 41—44.
О насильственном плаче при мозговых поражениях. Обозрение психиатрии, неврологии и экспериментальной психологии, 1906, № 11, стр. 742—749.
О некоторых рефлекторных явлениях.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 44—50.
О непроизвольном мочеиспускании при смехе.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1899, № 1, стр. 18—19.
О нервном симптомокомплсксе при простом ушибе, не осложненном так называемой «психической травмой».
Современная психоневрология,
1927, т. 4, № 1, стр. 2—9; № 2, стр. 116—126.
О парасимболии или транскорти-калыюй парафазии. Обозрение психиатрии, неврологии и экспериментальной психологии,
1909, № 9, стр. 513—517.
О подошвенной болезненной точке и подошвенно-пальцевом сгиба-телыюм рефлексе. Обозрение психиатрии, неврологии и экспериментальной психологии,
1910, № 7, стр. 403—405.
Реф. Обозрение психиатрии, неврологии и экспериментальной психологии, 1913, №-2, стр. 114—115.
О подлопаточном и грудном рефлексе. Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 12, стр. 899—901.
О подчревном рефлексе.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1900, № 2, стр. 81—82.
О поражении конского хвоста. Обо-
зрение психиатрии, неврологии и экспериментальной психологии, 1899, № 9, стр. 689—694.
О поражении конуса и хвоста спинного мозга fconus mcduilaris и caudae equinae). Врач, 1890, № 39, стр. 881—884.
О пояснично-бедренном рефлексе. Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, № 12, стр. 886—887.
О развитии женской груди у мужчин. Обозрение психиатрии, неврологии и экспериментальной психологии, 1907, № 3, стр. 186—187. То же. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905—1906 и 1906—1907 гг., СПБ, 1908, стр. 34—36.
О распознавательном значении феномена коленной чашки и других сродных явлениях. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 34—36.
О рефлексах в области лица и головы.
Обозрение психиатрии, неврологии и экспериментальной психологин, 1901, № 9, стр. 552—555.
О рефлексах дальних отделов верхней конечности и о диагностическом их значении. Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 6, стр. 407—410.
О рефлекторных явлениях при органических параличах центрального неврона и о новом тыльно-стопном сгибательном рефлексе ноги. Обозрение психиатрии, неврологии и экспериментальной психологии, 1911, № 7, стр. 385—389.
О своеобразном рефлекторном явлении, обнаруживаемом при насильственном подошвенном сгибании стопы и пальцев у больных с поражением центрального двигательного неврона. Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 8—9, стр. 771—772.
О сложных рефлекторных явлениях в нижних конечностях при поражениях центрального двигательного неврона. Обозрение психи-
495
атрии, неврологии и рефлексологии, 1927, № 1, стр. 1—16.
;) сложных сухожильно-мышечных рефлексах нижних конечностей при миэлитах.
Обозрение психиатрии, неврологии !! экспериментальной психологии, 1900, № 9, стр. 647—651.
О соотношении между обыкновенной и чувственной анестезией на основании клинических и экспериментальных данных. Неврологический вестник, 1893, т. 1, в. 2, стр. 3—20. ) сочетательно-двигательных рефлексах при церебральных параличах. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 127—129. Совместно с В. П. Протопоповым.
О транскортикальной парафазии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 9, стр. 666—676.
О фасциальных рефлексах нижних конечностей.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 12, стр. 889—892.
О феномене коленной чашки как распознавательном признаке нервных болезней и других сродных явлениях.
Обозрение психиатрии, неврологии и экспериментальной психологии. 1896, № 3, стр. 171 — 177. ) центральной гемиалгии, моналгии, термалгии и ригоралгии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 3, стр. 230. То же. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898-1899 гг., СПБ, 1899, стр. 24 — 26.
О частичных корковых и подкорковых параличах психорефлекторных ф>ункций.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1906, № 10, стр, 689—692.
О чувствительности к постукиванию скуловой дужки, как признаке органического поражения основных частей мозга и его оболочек. Обозрение психиатрии, неврологии
и экспериментальной психологи::, 1902, № 11, стр. 807—808.
Об акатизии.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902 — 1903 гг., СПБ, 1903, стр. 47—49.
Об акромиальном или верхушечном рефлексе.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1902, № 3, стр. 181 — 182.
Об извращенной световой реакции зрачков.
Неврологический вестник, 1897, т. 4, в. 2, стр. 165—171; в. 3, стр. 138—159.
Об изменениях мышечной чувствительности при спинной сухотке и других патологических состояниях.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 3, стр. 171 -172.
Об интенционном судорожном тике лица.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1913, № 4—5, стр. 193—198.
Об истощаемости сухожильных рефлексов и диагностическом их значении при нервных болезнях. Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 11, стр. 806—807.
Об одном своеобразном рефлекторном явлении, наблюдаемом в конечностях при центральных органических параличах. Обозрение психиатрии, неврологии и экспериментальной психологии,
1903, № 5, стр. 333—335.
Об особом лопаточно-плечевом рефлексе.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 10, стр. 767—768.
Об особом сгнбательном рефлексе ножных пальцев. Обозрение психиатрии, неврологии и экспериментальной психологии,
1904, Кя 6, стр. 421—422.
Об особом слуховом или акустико-па.тьпебральном рефлексе. Обозрение психиатрии, неврологии и экспериментальной психологии,
1905, № 2, стр. 86—88.
Об участии круговой мышцы глаза при корковых и подкорковых параличах лицевого нерва. Обо-
496
зрение психиатрии, неврологии и экспериментальной психологии,
1903, № 8, стр. 571—572. Офталмоплегия с периодическим поднятием и опусканием верхнего века и со своеобразной зрительной иллюзией.
Неврологический вестник, 1898, т. 6, в. 3, стр. 228—237; в. 4, стр. 36—69.
Пальцевый феномен на руке при органических гемиплегиях. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914 — 1915, № 10, 11 — 12, стр. 547—548.
По поводу пояснично-бедренного рефлекса.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1904, № 7, стр. 484—486. Показ больного с судорогой platysma
inyoides и некоторых шейных мышц. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901—• 1902 гг., СПБ, 1902, стр. 63—65.
Поражение варолиева моста с двусторонним параличом волевых движений глазных яблок, насильственным смехом и плачем и ранней атрофией мышц правой голени. Неврологический вестник, 1898, Т. 6, в. 4, стр. 160—212.
Приборы, служащие к точному клиническому исследованию коленных сухожильных рефлексов. Вестник клинической и судебной психиатрии и невропатологии, 1890, в. 1, стр. 43—53.
Pseudopolimelia paraesthetica в форме мнимых членов. Обозрение психиатрии, неврологии и рефлексологии, 1926, 4—5, стр. 236—241.
Pseudomelia paraesthetica как симптом черепно-мозгового поражения в области nucleus lenticularis. Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 12, стр. 896—900.
Распознавательное значение постукивания нижней части позвоночника. Неврологический вестник, 1897, № 3, стр. 6—7. То же. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—18К6 и 18*6—1897 гг., СПБ, 1898, стр. 42—43.
Симптомок омплекс нарушения нерв ных функций от воздействий, не осложненных так называемой «психической» травмой. В сб.: Новое в рефлексологии и физиологии нервной системы, в. 1, 1925, стр. 348—353. Автореферат совместно с Г. Е. Шум-ковым.
Случай надглазной атрофии.
Протоколы заседаний общества психиатров за 1906—1907 гг., СПБ, 1907, стр. 64.
Случай судороги в плечевом поясе в виде маятникообразного движения то одного, то другого плеча. Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 8, стр. 620—621.
Тахифемия, тахибазия и пароксиз-мальное топтание на месте как постэнцефалитический синдром. Обозрение психиатрии, неврологии и экспериментальной психологии, 1924, № 1, стр. 24—28.
Электрический трихоэстемометр для исследования так называемой во-лосковой чувствительности тела. Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 10, стр. 777—779.
Выступления в прениях по поводу сообщений
Анфимов В. Я., Демонстрация больного (спинная сухотка и атрофия конечностей). Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней г,а 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 132.
Бари А. Э., Демонстрация больной с викариирующим менструальным кровотечением. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 26—27.
Бари А. Э. и Карпинский А. И., Демонстрация больной с мозговой аневризмой. Протоколы заседаний Общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр. 43—44.
Б л у м е н а у Л. В., К вопросу о самородном дрожании. Отчеты научных собраний врачей С.-Петербургской клиники душев-
32 В. М. Бехтереи
497
ных и кговных болезней за 1901 — 1902 гг.,'СПБ, 1902, стр. 27—28.
Бух М. Г., О невралгиях симпатического нерва и о болезненной раздражительности его вообще с преимущественным рассмотрением болезненных ощущений в брюшной и тазовой полости и их отношение к симпатическому и блуждающему нерву.
Протоколы заседаний Общества психиатров за 1900 и 1901 гг., СПБ, 1902, стр. 28.
Гольцингер Ф. Ф., О ладонном рефлексе.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 62.
Гольцингер Ф. Ф., Распознавательное значение ладонного рефлекса.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 33—34.
Грибоедов А. С. и Бруш-т е й н А. Н., О значении кожно-пальцевых рефле'ксов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900 — 1901 гг., СПБ, 1901, стр. 66—67.
Данилло С. Н., О различных видах дрожания.
Протоколы заседаний Общества психиатров за 1892 г., СПБ, 1893, стр. 26.
Жуков Н. А., Исследования звукопроводимости костей черепа и позвоночника электрическим камертоном и его распознавательное значение.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 12.
Жуковский М. Н., О подош-венно-пальцевом сгибательном феномене.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908'— 1909 и 1909—1910 гг., СПБ, 1910, стр. 118—119.
Жуковский М. Н., Случай хейромегалии, развившейся вслед за травмой у больного, страдающего сирингомиэлией.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ, 1908, стр. 63—64.
Иванов И., О макроэстезии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 89—S0.
Карпинский А. И., Демонстрация больной с нарколептиче-скими явлениями на почве соп-stitutio thyreotoxice. Протоколы заседаний Общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр. 90—91.
Карпинский А. И., Демонстрация больной, страдающей болезнью Dercum'a (adipositas do-lorose).
Протоколы заседаний Общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 19.
Кульбин Н. Н., Демонстрация нового прибора для исследования тактильной и болевой чувствительности.
Протоколы заседаний Общества психиатров за 1894 г., СПБ, 1896, стр. 17—18.
Ларионов В. Е., Два случая транскортикальной чувственной и двигательной афазии с сохранением музыкальных способностей. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 28.
Никитин М. П., К. симптоматологии поражений зрительного бугра.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1910— 1911 гг., СПБ, 1911, стр. 35—36.
Никитин М. П., Клонус стопы функционального происхождения. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1910— 1911 гг., СПБ, 1911, стр. 5—6.
Ноишевскии К. И., О кортикальной гемиопии. Протоколы заседаний Общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 59—60.
Ноишевскии К- В., Отношение между застойным соском и экскавацией зрительного нерва.
498
Протоколы заседаний Общества психиатров за 1909 и 1910 гг. СПБ, 1911, стр. 47. Ольдерогге В. В., Демонстрация больного.
Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ,
1897, стр. 89—90. Осипов В. П., Демонстрация
больного, страдающего сальтатор-ной судорогой.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 —
1902 гг. СПБ, 1902, стр. 55— 56.
Осипов В. П., Значение рефлекса Бабинского при органических и функциональных геми-плегиях.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902—
1903 гг., СПБ, 1903, стр. 116—118. Останков П. А., О кожных
рефлексах в начальном периоде tabes'a.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ,
1898, стр. 91—92. Останков П. А., О периодической рвоте табетиков (гастрические кризы).
Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 64—65.
П у с с е п Л. М., Об остром половинном отеке при гемиплегии. Отчеты научных собраний врачей С.-Петербургской клиники душев-
, ных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ, 1908, стр. 38.
С н и к е р П. М., Демонстрация больного с расстройством стерео-гностической чувствительности. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902— 1903 гг., СПБ, 1903, стр. 8—9.
Фальк М. Л. Демонстрация больного.
Протоколы заседаний Общества психиатров за 1900 и 1901 гг., СПБ, 1902, стр. 43—50.
Франк Д. Б., О расстройстве зрения при множественном склерозе.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 78—79. Чудновский Я. Б., Об изменениях чувствительности при спинной сухотке (tabes dorsalis) с обращением особенного внимания на волосковую и мышечную чувствительность.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 80—82.
Нервные болезни
Книги
Нервные болезни в отдельных наблюдениях.
в. 1—2, Казань, 1894—1899 гг., в. 1, 255 стр., в. 2, 278 стр.
Общая диагностика болезней нервт ной системы,
часть 1—2, СПБ, 1911—1915, 332 стр.
Сифилис центральной нервной системы, СПБ, 1902, 116 стр.
Статьи
Апоплектическая гемитония. Обозрение психиатрии, неврологии и экспериментальной психологии,
1899, № 7, стр. 540—547, № 8, стр. G31—637.
Болезненная неподвижность (akinesia algera).
Неврологический вестник, 1893, т. 1, в. 2, стр. 99—146.
Гемихорея как результат кровоизлияния в thalamus opticus. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 58—59.
Два случая сифилитического поражения мозга.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1900, № 10, стр. 783. Демонстрация больного с поражением задней половины спинного мозга.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907— 1908 гг., СПБ, 1909, стр. 13—16. Демонстрация больных (гемитония). Отчеты научных собраний врачей
32* Щ,
С.-Петербургской клиники душевных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ, 1908, стр. 64—66. Демонстрация больного с афазией. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1904—
1905 гг., СПБ, 1905, стр. 1—4. Демонстрация больных (ранение черепа и др.).
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905—
1906 и 1906—1907 гг., СПБ, 1908, стр. 21—24.
Демонстрация больного, только что оправившегося от tetanus'a. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 9—10.
Демонстрация tumor cerebri.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 72—73. К этиологии и клинике «одервене-лости» позвоночника. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915, № 7—9, стр. 467—474.
Leptomeningitis haemorrhagica, протекавший с явлениями прогрессивного паралича помешанных. Еженедельная клиническая газета, 1881, № 8, стр. 138—141; 1881, № 9, стр. 153—157; № 10, стр. 170—176.
Macrocephalia resp. macromelia paraesthetica.
Современная психоневрология, 1928, № 2, стр. 121 — 130. Совместно с Я. А. Ратнером.
Миотония как болезнь обмена. Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 4, стр. 263—268.
Нервное слюнотечение как самостоятельное болезненное расстройство. Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 2, стр. 92—97.
Новый случай одеревенелости позвоночного столба с патологоанато-мическими исследованиями и об анкилозирующем воспалении позвоночника и больших суставов. Отчеты научных собраний врачей
С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 42—45.
О болезненной форме, описанной в последнее время Мебиусом под названием «akinesia alegra». Неврологический вестник, 1894, т. 2, в. 1, стр. 53—54.
О миопатической гипертрофии мышц. Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 3, стр. 176—185.
О неврозе в форме интеиционных тонических судорог. Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 1, стр. 14—22.
О нервной форме поражения с внешним характером миотонии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 10, стр. 744—751.
О плясковой падучей.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ,
1898, стр. 45—47.
О поражении мозговой коры при рассеянном склерозе. Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 7, стр. 540.
О рефлекторной эпилепсии под влиянием звуковых раздражений. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915, № 10, 11, 12, стр. 513— 520.
О сифилитическом рассеянном че-репноспинном склерозе. Обозрение психиатрии, неврологии и экспериментальной психологии,
1896, № 4, стр. 255—265; № 5, стр. 342—352; № 6, стр. 413—418.
О сращении или одеревенелости позвоночного столба как особой форме заболевания.
Неврологический вестник, 1897, т. 5, в. 1, стр. 147—158.
О хореической падучей. Обозрение психиатрии, неврологии и экспериментальной психологии,
1897, № 10, стр. 728—733.
Об анкилозирующем воспалении позвоночника и больших суставов конечностей.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1899, № 3, стр. 198—206.
500
Об одеревенелости или сращении позвоночника с нервным симптомо-комплексом.
Русский врач, 1913, № 30, стр. 1053—1056; № 31, стр. 1085—1087.
Об особом, развивающемся с детства, нервном поражении, выражающемся двигательными расстройствами и слабоумием.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, № 3, стр. 162—169.
Об острой мозжечковой атаксии. Обозрение психиатрии, неврологии и экспериментальной психологии,
1901, № 6, стр. 413—416. Одеревенелость позвоночника с искривлением его как особая форма заболевания.
Врач, 1892, № 36, стр. 899—903. Остро развивающееся расстройство
движения у пьяниц с характером
мозжечковой атаксии.
Обозрение психиатрии, неврологии
и экспериментальной психологии,
1900, № 1, стр. 1—4. Paramyoclonus multiplex.
Врач, 1887, № 3, стр. 77—78;
№ 5, стр. 99—101; № 6, стр. 135—
137. Случай акроэритрозиса (Асгоегу-
trosis).
Протоколы заседаний Общества
психиатров за 1906—1907 гг., СПБ,
1907, стр 64—65. Случай опухоли мозжечка.
Обозрение психиатрии, неврологии
и экспериментальной психологии,
1902, № 3, стр. 226. Смертельно протекшая пляска беременной женщины.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 19—20.
Тетания.
Неврологический вестник, 1894,
т. 2, в. 2, стр. 137—166. Тяжкая пляска (Chorea gravis).
Неврологический вестник, 1896,
т. 4, в. 3, стр. 39—78.
Hemiatrophia facialis progressiva. Вестник клинической и судебной психиатрии и невропатологии, 1888, в. 1, стр. 228—235.
Выступления в прениях по поводу сообщений
Антушевич В. И., О миокло-нии.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903— 1904 гг., СПБ, 1904, стр. 15—17.
Аствацатуров М. И., О рефлекторных явлениях в речевой сфере.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907—•
1908 гг., СПБ, 1909, стр. 7—8. Бари А. Э., Демонстрация больного с diplegia spastica cerebralis. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898—-1899 гг., СПБ, 1899, стр. 55.
Бари А. Э., По поводу одного случая периферического паралича п. facialis.
Протоколы заседаний Общества психиатров за 1898 г., СПБ, 1899, стр. 32.
Б и р у л я-Б ялыницкий Ф.А., Arachnitis spinalis ossificans. Протоколы заседаний общества психиатров за 1900—1901 гг., СПБ, 1902, стр. 21—22.
Б и р у л я-Б ялыницкий Ф. А., Случай цистицерков мозга. Протоколы заседания общества психиатров за 1894 г., СПБ, 1896, стр. 4—5.
В а л и ц к а я М. К., Демонстрация больной (случай лицевого паралича).
Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 91.
Вендерович Е. Л., Поражение каудально-вентрального отдела thalami.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—
1909 и 1909—1910 гг., СПБ, 1910, стр. 102—103.
Вырубов Н. А., Три случая мозговых опухолей с предъявлением препаратов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 63.
501
Гольцингер Ф. Ф., Латиризм в Абиссинии.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 97—98.
Гольцингер Ф. Ф., Случай кессонной параплегии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900— 1901 гг., СПБ, 1901, стр. 14—15.
Горловский А. Д., Демонстрация больного с одеревенелостью позвоночника.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ, 1908, стр. 54, 55—56.
Жуков Н. А., О клинических наблюдениях над звукопроводимостью костей черепа и позвоночника при помощи электрического камертона.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 16—17.
Иванов И. И., К симптоматологии и патологической анатомии амиотрофического бокового склероза.
Протоколы заседаний Общества психиатров за 1902—1903 гг., СПБ, 1904, стр. 41.
Иванов И. И., Случай скрытого течения мозговой опухоли. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902—
1903 гг., СПБ, 1903, стр. 30—31. Корольков П. Я., Демонстрация ребенка, пораженного слизистым отеком.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903—
1904 гг., СПБ, 1904, стр. 90—91. Корольков П. Я., Демонстрация ребенка с параличом шейных мышц.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 46.
502
Кудрявский Г. Н., Демон* страция больного (случай морва-новой болезни).
Протоколы заседаний общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 52.
Любимов С. Я., Множественные опухоли мозга.
Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 20—21.
Неаронов В. М., Демонстрация случая миотонии. Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 80—81.
Никитин М. П., Случай атрофии в области разветвления 1-й ветви тройничного нерва. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—■ 1909 и 1909—1910 гг., СПБ, 1910, стр. 65—66.
Никитин М. П., Случай паралича п. cruralis и п. obturatorii. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907—
1908 гг., СПБ, 1909, стр. 5—6. Осипов В. П., Демонстрация
больного с одеревенелостью позвоночника.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 82—83. П у с с е п Л. М., Демонстрация больного с шейным ребром. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—
1909 и 1909—1910 гг., СПБ, 1910, стр. 114.
П у с с е п Л. М., О нервном ограниченном отеке.
Протоколы заседаний Общества психиатров за 1906—1907 гг., СПБ,
1907, стр. 46—47.
П у с с е п Л. М., Случай родового паралича.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ,
1908, стр. 61.
Р е й м а н Н. К., Два случая одеревенелости позвоночника. Прото-
колы заседаний Общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 80—81.
Р о з е н б а х П. Я-, О патогенезе падучей болезни. Протоколы заседаний Общества психиатров за 1883 г., СПБ, 1885, стр. 32—34.
Р ы б а л к и н Я- В., Демонстрация больного.
Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 99—100.
Рыбалки н Я. В., Демонстрация трех нервнобольных. Протоколы заседаний Общества психиатров за 1899 г., СПБ, 1901, стр. 23—26.
Скоп В. В., Случай болезни Raynaud.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 33.
Спиртов И. Н., К вопросу о Brown-Sequard'oBCKoft эпилепсии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905— 1906 и 1906—1907 гг., СПБ, 1908, стр. 28—29.
Т р о ш и н Г. Я-, Случай одеревенелости позвоночника. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902— 1903 гг., СПБ, стр. 39—40.
Трошин Г. Я- и Пуссеп Л.М., К казуистике одеревенелости позвоночника.
Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902— 1903 гг., СПБ, 1903, стр. 56—57.
Методы исследования и общая
симптоматика психических
заболеваний
Книги
Записки по психиатрии по лекциям проф. Бехтерева, 1907—1908 гг. Составлен слушательницами V курса женского медицинского института Залесской и Соловцевой, СПБ, 1907, 210 стр.
Опыт клинического исследования температуры тела при некоторых формах душевных заболеваний.
Диссертация на степень доктора медицины. Военно-мед. журнал. 1880, 4, 139, отдел II, стр. ИЗ— 168; 1881, 4, 140, отдел II, стр. 41 — 120. / Тоже, отд. изд., СПБ, 1881,315 стр.
Статьи
Автоматизм как одно из проявлений мозгового сифилиса. Обозрение психиатрии, невропатологии и экспериментальной психологии, 1896, № 9, стр. 662—665.
Апоплекто- и эпилептовидные припадки в прогрессивном параличе помешанных.
Международная клиника, 1882, № 8, стр. 1—24 (549—554). Реф. Протоколы Общества психиатров в С.-Петербурге, 1881, СПБ, 1883, стр. 20—23;
Боязнь покраснеть. Обозрение, 1897, № 1, стр. 23—24.
Боязнь полового бессилия и coitus'a как причина половой неспособности.
Обозрение психиатрии, невропатологии и экспериментальной психологии, 1907, № 2, стр. 85—98.
Боязнь чужого взгляда. Русский врач, 1905, № 3, стр. 73—77.
Взаимовнушение в сообществе людей и коллективные галлюцинации. Вестник знания, 1926, № 6, стр. 361—366.
Вильгельм — дегенерат нероновско-го типа.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915, № 7, 9, стр. 399—418.
Внушение как фактор преступления (по поводу окончательного судебного решения по делу Румянцевой). Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898— 1899 гг., СПБ, 1899, стр. 83—86.
Дальнейшие наблюдения над «боязнью покраснеть». Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 5, стр. 323—329.
Демонстрация больного с расстройством в отношении чувства направлений, развившегося под влиянием травматического шока. Протоколы заседаний Общества
503
психиатров в С.-Петербурге за 1912 г., СПБ, 1912, стр. 13—14.
Демонстрация больного со своеобразным расстройством речи, развившимся в ввязи с функциональным неврозом.
Протоколы заседаний Общества психиатров в С.-Петербурге за 1912 г., СПБ, 1912, стр. 11 — 13.
Инспираторные вздохи у истеричной. Обозрение психиатрии, неврологии и экспериментальной психологии, 1907, № 8, стр. 508—509.
К вопросу о записывающем способе исследования сухожильных рефлексов и об изменении их при душевных болезнях. Врач, 1891, № 16, стр. 397—399.
Кликушество и насильственные крики как симптом истерии. Обозрение психиатрии, неврологии и экспериментальной психологии,
1899, № 11, стр. 920—921. Краснение лица как особый вид
болезненного расстройства. Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 12, стр. 899—903.
Материалы к методике объективного исследования душевнобольных. Вестник психологии, криминальной антропологии и гипнотизма, 1910, № 5, стр. 386—430. Совместно с С. Д. Владичко.
Насильственные автоматические движения и письмо как симптомы душевного расстройства. Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 6, стр. 412—422.
Насильственное произношение слов в виде самостоятельного болезненного расстройства. Неврологический вестник, 1897, № 3, стр. 3. Реф. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 и 1896— 1897 гг., СПБ, 1898, стр. 39—40.
Невроз местной контузии.
Новое в рефлексологии и физиологии нервной системы, Л. •— М., 1925, в. 1, стр. 344—347. Совместно с Г. Е. Шумковым.
Непереносливость или боязнь чужого взгляда.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1900, № 7, стр. 491—497.
Новые наблюдения над боязнью покраснеть.
Обозрения психиатрии, неврологии и экспериментальной психологии, 1897, № 8, стр. 561—566.
О болезненном страхе профессионального характера.
Боязнь великого выхода у священников.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 10, стр. 733—735.
О болезненных ощущениях и соощу-щениях у душевных больных. Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 2, стр. 103—112.
О больных, преследуемых запахами. Обозрение психиатрии, неврологии и экспериментальной психологии, 1906, № 1, стр. 63.
О боязни опоздать.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1906, № 7, стр. 478—481.
О бреде гипнотического очарования. Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 4, стр. 249—263.
О внушающем влиянии слуховых обманов чувств.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № 11, стр. 822—824.
О внешних признаках привычного ' онанизма у подростков мужского пола.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 3, стр. 658—663.
О галлюцинаторных воспоминаниях. Обозрение психиатрии, неврологии и экспериментальной психологии,
1907, № 6, стр. 328—333.
О значении внимания в отношении локализации и развития галлюцинаторных образов. Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 11, стр. 825—832.
О навязчивой рвоте. Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 6, стр. 419—422.
О навязчивой ревности.
Русский врач, 1908, № 34, стр. 1121 — 1124.
О навязчивой улыбке. Обозрение психиатрии, неврологии
504
и экспериментальной психологии, 1899, № 6, стр. 449—452.
О навязчивом потении рук. Обозрение психиатрии, неврологии и 'экспериментальной психологии, 1905, № 7, стр. 485—486.
О навязчивых кишечных и пузырных кризах и о значении при них лечения внушением. Обозрение психиатрии, неврологии и экспериментальной психологии, 1907, № 1, стр. 10—17.
О навязчивых ненормальных ощущениях и навязчивых обманах чувств.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1899, № 2, стр. 87—96.
О навязчивых фобиях и их лечении. Русский врач, 1915, № 14, стр. 313—316.
О нарушении обмена веществ при неврастении.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 7, стр. 527—529.
О нарушении чувства времени у душевнобольных.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 3, стр. 181 — 185.
О периодических приступах ретроактивной амнезии. Обозрение психиатрии, неврологии и экспериментальной психологии,
1900, № 4, стр. 248—253.
О половых извращениях как патологических сочетательных рефлексах.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915, № 7, 9, стр. 357—382.
О психических актах глотания (dysphagia psychica).
Обозрение психиатрии, неврологии и экспериментальной психологии,
1901, № 5, стр. 321—324.
О расстройствах в сфере чувственного восприятия у душевнобольных.
Обозрение психиатрии, неврологии и экспериментальной психологии,
1902, № 4, стр. 245—253.
О ритмических судорогах и автоматических движениях истеричных. Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, № 4, стр. 250—255.
О своеобразном психопатическом затруднении мочеиспускания. Обо-
зрение психиатрии, неврологии и экспериментальной психологии, 1897, № 12, стр. 884—891.
О слышании собственных мыслей. Обозрение психиатрии, неврологии и экспериментальной психологии, 1896, № ю, стр. 733—740.
О состоянии мышечных и других рефлексов лица при нарастающем параличном слабоумии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 4, стр. 244—246.
О термических изменениях у душевнобольных.
Медицинский вестник, 1879, № 7, стр. 52—54; № 8, стр. 58—60; № 9, стр. 65—67; № 10, стр. 74—76; № 13, стр. 102, 104; № 14, стр. ПО—112; № 15, стр. 118—120; № 17, стр. 135; № 18, стр. 142— 144; № 20, стр. 159—160; № 21, стр. 167—169
Об искусственном вызывании обманов чувств у алкоголиков, страдающих галлюцинаторными формами помешательства. Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 2, стр. 101 — 104.
Объективное исследование душевнобольных.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1907, № 10, стр. 595—606; № 11, стр. 641—647; № 12, стр. 705—716.
Об объективных признаках местной гиперестезии и анестезии, наблюдаемых при так называемых травматических неврозах и при истерии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 11, стр. 865—868.
Об объективных признаках расстройств чувствительности при так называемых травматических неврозах.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, № 1, стр. 19—26.
Об одной своеобразной форме фетишизма.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 1, стр. 11 — 13.
Об экспериментально-объективном исследовании душевнобольных. Обозрение психиатрии, неврологии и экспериментальной психологии, 1910, № 10—11, стр. 648—664.
505
Объективное исследование больной личности как основа патологической рефлексологии. В сб. Научная медицина, № 9, Пг, 1922, стр. 1—9.
Объективное исследование душевнобольных.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1907, № 10, стр. 595—606; № 11, стр. 641—647; № 12, стр. 705—716.
Одержимость гадами. Обозрение психиатрии, неврологии и экспериментальной психологии, 1900, № 5, стр. 330—332.
Половы» извращения и уклонения в свете рефлексологии. В кн.: Вопросы изучения и воспитания личности, Пг., 1922, в. 4—5, стр. 644—746.
Предисловие к кн.: Н. В. Краинский. Порча, кликушество и бесноватые как явления русской народной жизни, Новгород, 1900, 243 стр.
Признание действительности за галлюцинаторное переживание как одно из проявлений психического расстройства.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1906, № 6, стр. 413—418.
Случай полной слепоты, сопровождаемый галлюцинациями зрения. Протоколы заседаний Общества психиатров в С.-Петербурге за 1883, СПБ, 1885, стр. 54—57.
Состояние оцепенелости при melancholia attonita. Международная клиника, 1882, № 4, стр. 1—23. Лекция читана в конференции Военно-медицинской академии на звание приват-доцента.
Эпилептические и эпилептоидные приступы, протекающие под видом страха.
Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 12, стр. 920—927.
Выступления в прениях по поводу сообщений
Абрамов В. В., Экспериментально-психологические исследования больных с разными формами слабоумия и больных с маниакально-меланхолическим психозом. Протоколы заседаний общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр.65—66.
Б л у м е н а у Л. В., Атаксия вследствие свинцового отравления (от дурной полуды). Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 53.
БлуменауЛ. В., К патологии полового влечения. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за- 1901 — 1902 гг., СПБ, 1902, стр. 23—25.
Б л у м е н а у Л. В., Случай частичного пареза прямых мышц живота у истеричного субъекта. Отчеты научных собраний вра-* чей С.-Петербургской клиники душевных и нервных болезней за 1897—1898 гг., СПБ, 1899, стр. 49—50.
Б р а ж а с В. Н., Демонстрация нервнобольной с парезом диафрагмы. Протоколы заседаний Общества психиатров за 1909 и ' 1910 гг., СПБ, 1911, стр. 16.
Владимирский А. В., Истерия душевнобольного с религиозным бредом во время прошлой войны. Протоколы заседаний общества психиатров за 1906— 1907 гг., СПБ, 1907, стр. 70.
Воскресенский М. К..> Случай серпигинозной гангрены ко- . ■ жи у душевнобольной. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899—1900 гг., СПБ, 1900, стр. 62.
Гутман Л. П., Состояние сосредоточения и умственная работоспособность при маниакально-депрессивном психозе. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—1909 и 1909—1910 гг., СПБ', 1910, стр. 83—84.
Добротворский В. И., Демонстрация больного с амне-зиею за весь период психического расстройства. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 48—49.
Завадский Н. К., Характер ассоциаций у больных с рага-
506
noia chronica. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908— 1909 и 1909—1910 гг., СПБ, 1910, стр. 47—48.
Иванов И. И., К вопросу о половом извращении. Протоколы заседаний общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 97.
Корольков П. Я-, Случай спастической диплегии у ребенка. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 53—54.
К р а и н с к и й Н. В., Кликушество, порча и беснования в России. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899—1900 гг., СПБ, 1900, стр. 16—17.
К р а и н с к и й Н. В., Новые наблюдения эпидемии кликушества в Новгородской губернии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899—1900 гг., СПБ, 1900, стр. 55—56.
Никитин М. П., О массовых галлюцинациях и иллюзиях. Протоколы заседаний общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 114.
П^у с с е п Л. М., Редкий случай arthropatiae при прогрессивном параличе помешанных. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900 гг., СПБ, 1900, стр. 60.
Р ад и н Е. П., Истерический сколиоз на почве одностороннего сведения мускулатуры спины. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905—1906 и 1906—1907 гг., СПБ, 1908, стр. 24—25.
Р а д и н Е. П., Методика психологического исследования бреда и краткий обзор учений о бреде. Вестник психологии,криминальной антропологии и гипнотизма, 1904 № 1, стр. 83.
Р а д и н Е. П., Об извращенном
симвалически - эгоцентрическом восприятии окружающего в состоянии концентрации при паранойе, при параноидных состояниях. Протоколы заседаний общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 64— 65.
Резников X. М., О поле зрения у больных, страдающих прогрессивным параличом помешанных. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897—1898 гг., СПБ, 1899, стр. 58.
С е м е к а Д. В., К казуистике острых сыпей у душевнобольных. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897—1898 гг., СПБ, 1899, стр. 61—62.
Сокальский Н. А., Иннервация сосудов при острых душевных болезнях. Протоколы заседаний общества психиатров за 1899 г., СПБ, 1901, стр. 40—41.
Спиртов И. Н., К вопросу о кровяном давлении у душевнобольных. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905—1906 и 1906— 1907 гг., СПБ, 1908, стр. 74—75.
Юрман Н. А., Случай трихотил-ломании. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900—1901 гг., СПБ, 1901, стр. 23.
Юрман Н. А., Случай травматического невроза. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ, 1898, стр. 60—61.
Психические болезни Книги
Классификация душевных болезней. Ученые записки Казанского университета, Казань, 1891; то же, отд. изд., Казань, 1891, 60 стр.
Неврозы и психоневрозы. В кн.: Частная патология и терапия внутренних болезней. Под ред. Г. Ф. Ланга. М.—Л., 1929, т. 4,
507
в. 3, стр. 807—938. Совместно с Р. Я. Голант. Психопатия (психо-нервная раздражительная слабость) н ее отношение к вопросу о вменении. Дн. Казанского общества врачей при университете, 1886, № 2—3, стр. 10—40, то же, отд. изд., Казань, 1886, 31 стр. Реф. Архив психиатрии, неврологии и судебной психопатологии, 1885, т. 7, № 3, стр. 75.
Статьи
Бред гипнотического очарования или «paranoia suggestia deliria». Русский врач, 1913, № 4, стр. 111 — 113.
Демонстрация душевнобольной, страдавшей чувственным бредом. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902—1903 гг., СПБ, 1903, стр. 1—6.
К учению о микроцефалии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 5, стр. 343—348. Совместно с, М. Н. Жуковским.
О галлюцинаторном психозе, развивающемся при поражении органа слуха. Обозрение психиатрии, неврологии и экспериментальной психологин, 1903, № 2, стр. 91 — 106.
О маниакально-меланхолическом психозе. Обозрение психиатрии, неврологии и экспериментальной психологии, 1909, № 11, стр. 641—657. Реф. Обозрение психиатрии, неврологии и экспериментальной психологии, 1909, № 3, стр. 188—189.
О периодической острой паранойе как особом виде периодических психозов. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 4, стр. 279—293.
О своеобразной нервно-психической эпидемии среди рабочих рижских и петроградских фабрик, бывшей в марте 1914 г. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915, № 7—9, стр. 419— 466; № 10—12, стр. 585—594.
О своеобразном психозе, развивающемся на почве хронического
поражения обонятельного органа (паросмофоння). Обозрение психиатрии, неврологии и экспериментальной психологии 1906 № 2, стр. 102—113.
О соотношении между маниакальными и меланхолическими состояниями. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—1909 и 1909—1910 гг., СПБ, 1910, стр. 3—7.
Об извращении и уклонении полового влечения с рефлексологической точки зрения. В сб.: Половой вопрос в свете научного знания, Госиздат, 1926, стр. -293—325.
Помешательство с бредовым внушением Paranoia suddestio deliria. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914, № 4—6, стр. 209—224.
Соматофрения. Обозрение психиатрии, неврологии и рефлексологии, 1928, № 1, стр. 5—12.
Убийство должностного лица душевнобольным, страдающим paranoia chronica. Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 9, стр. 695—701; № 10, стр. 747 — 761; К« 11, стр. 843—854.
Выступления в прениях по поводу сообщений
Агаджаняиц К- С., Есть ли первичное острое сумасшествие самостоятельная нозологическая единица. Протоколы заседаний Общества психиатров за 1902 н 1903 гг., СПБ, 1904, стр. 74.
Агаджаняиц К- С, Индуцированное помешательство и психическая зараза. Протоколы заседаний Общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 95.
Б л у м е н а у Л. В., О проявлениях дегенеративной слабости нервной системы у лиц простого звания (по вопросу об истерии у солдат). Протоколы заседаний Общества психиатров за 1898 г., СПБ, 1899, стр. 39.
Б л у м е н а у Л. В., Случай микроцефалии. Отчеты научных со-
508
браний врачей С.-Петербургской клиники душевных и нервных болезней за 1897—1898 гг., СПБ, 1899, стр. 48.
Гутман Л., Маниакально-депрессивный психоз и характер ассоциации прн нем. Протоколы заседаний Общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр. 39—41.
Игнатьев М. В., Данные статистики об идиотизме и imbecil-1 itas. Протоколы заседаний Общества психиатров за 1895— 1896 гг., СПБ, 1897, стр. 22—23.
Коссаковский В. Л., О пеллагре, наблюдавшейся в 1888 г. в психиатрическом отделении губ. Бессарабской земской больнице. Протоколы заседаний общества психиатров за 1894 г., СПБ, 1896, стр. 12—13.
Лебедев И. П., Случай травматического повреждения черепа. Протоколы заседаний Общества психиатров за 1899 г., СПБ, 1901, стр. 14—15.
Никитин М. П., К вопросу о кликушестве. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903—1904 гг., СПБ, 1904, стр. 5—6.
Ольдерогге В. В., Отсутствие мышечного труда у душевнобольных как этиологический момент легочных заболеваний. Протоколы заседаний Общества психиатров за 1894 г., СПБ, 1896, стр. 37—38.
Павловская Л. С, К вопросу о первичном сумасшествии как об особой клинической форме в связи с вопросом о причинах изменения классификации психозов.
Суханов С. А., Существует ли паранойя как самостоятельная болезнь. Протоколы заседаний Общества психиатров за 1909—1910 гг., СПБ, 1911, стр. 59—60.
Р а д и н Е. П., Переходные формы между истерией и истерическим психозом. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903—1904 гг., СПБ, 1904, стр. 69—70.
Рейтц Г. В., Случай dementia
praecox. Протоколы заседаний Общества психиатров за 1900— 1901 гг., СПБ, 1902, стр. 59.
Скворцов К- Н-, Опухоль мозжечка у душевнобольной. Протоколы заседаний Общества психиатров за 1894 г., СПБ, 1896, стр. 35.
Сол у к И. П., Преступление, совершенное в состоянии патологического опьянения. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900—-1901 гг., СПБ, 1901, стр. 35—36.
Суханов С. А., Циклотимия и маниакально-депрессивный психоз. Протоколы заседаний Общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр. 19—20.
Фи нкель штейн Л. О., Два случая folie transformer. Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 78.
Ю щ е н к о А. И., Об окислительном процессе в организме душевнобольных и ядовитости мочи у них. Протоколы заседаний Общества психиатров за 1906— 1907 гг., СПБ, 1907, стр. 59—60.
Ю щ е н к о А. И., О рефлекторных центрах в узлах симпатической нервной системы, об отношении нижнего брыжеечного узла к иннервации мочевого пузыря и of) автоматических движениях последнего. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895 г. и 1896— 1897 гг. СПБ, стр. 96—97.
Лечение нервных и психических заболеваний
Книги
Гипноз, внушение и психотерапия и их лечебное значение, СПБ, 1911, 60 стр.
К вопросу о врачебном значении гипноза. Неврологический вестник, 1893, т. 1, в. ], стр. 27—44. То же, отд. изд., Казань, 1893, 20 стр.
Лечебное значение гипноза. Спутник здоровья, 1900, № 1, стр. 2—3; № 2, стр. 23—25; № 4, стр. Q2— 63; № 7, стр. 122—123; № 9, стр. 160—161; № 12, стр. 220—
509
221; № 15, стр. 280—281. То же, отд. изд., СПБ, 1900, 35 стр.
Статьи
Adonis vernalis и препарат его Ado-nilen в лечении эпилепсии и общих неврозов. Клиническая медицина, 1925, № 6 (34), стр. 172— 173. Совместно с Г. А. Певзнер.
Внушение и чудесные исцеления. Вестник знания, 1925, № 5, стр. 321—332.
Вопросы, связанные с лечебным и гигиеническим значением музыки. Обозрение психиатрии, неврологии и экспериментальной психологии, 1916, №1—3, стр. 105— 124.
Гипноз. Вестник знания, 1925, № 16, стр. 1057—1064.
Гипноз и значение его как врачебного средства. Неврологический вестник, 1894,т. 2, в. 3, стр.1—58.
Значение подвешиваний при некоторых спинномозговых поражениях. В кн.: Б. И. Воротынский, Подвешивание как метод лечения нервных болезней. Казань, 1893, стр. 65—72.
Лечение внушением превратных половых влечений и онанизма. Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 8, стр. 587— 597.
Навязчивые кишечные и пузырные кризы, их лечение внушением. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905, 1906 и 1906—1907 гг., СПБ, 1908, стр. 39—42.
О значении сердечных средств в ле-черии падучей. Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 6, стр. 401—406.
О значении смеси из Adonis vernalis или Digitalis с бромидами и кодеином в лечении падучей. Обозрение психиатрии, неврологии И экспериментальной психологии, 1898, N° 9, стр. 679— 681. О значении совместного употребления бромидов Adonis vernalis .при падучей. Неврологический вестник, 1894, т. 2, в. 3, стр.101 — 108.
О значении совместного применения гипнотических внушений и других средств при лечении привычного пьянства. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 1, стр. 8—10. О лечебном значении умственного труда для больных общими неврозами. Вопросы психофизиологии, рефлексологии и гигиены труда, сб. 1, Казань, 1923, стр. 5—12. О лечении болезненных влечений и навязчивых состояний отвлекающей психотерапией. Обозрение психиатрии,неврологии и экспериментальной психологии, 1913, № 6—7, стр. 321—329. О лечении виттовой пляски. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 12, стр. 937—941. О лечении врачебной гимнастикой в ваннах. Обозрение психиатрии, неврологии и экспериментальной психологии, 1903, № 9, стр. 649—650. О лечении гипнозом. Вестник знания, 1926, № 2, стр. 85—94. О лечении миотонии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 5, стр. 321—334. О лечении навязчивых идей гипнотическим внушением, Врач, 1892, № 1, стр. 1—2. О лечении онанизма внушениями в гипнозе. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 3, стр. 186—189. О миотонии и ее лечении. Неврологический вестник, 1896, т. 4, в. 4, стр. 107—130. О психорефлекторной терапии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914—1915, № 10, 11, 12, стр. 555—557. О сочетательно-рефлекторной терапии. Вестник современной медицины, 1925, № 11, стр. 2—4. О терапевтическом значении воспитания сочетательных рефлексов при истерических анестезиях и параличах. Обозрение психиатрии, неврологии и рефлексологии, 1917—1918, № 1—2, стр. 54—55. Об объективных признаках внушений, испытываемых в гипнозе. Вест-
510
ник психологии, криминальной антропологии и гипнотизма, 1905, № 4, стр. 275—280.
Об оперативном вмешательстве при хореической падучей. Обозрение психиатрии, неврологии и экспериментальной психологии, 1901, № 7, стр. 487—492.
Объективные признаки внушенных изменений чувствительности в гипнозе. Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 1, стр. 12— 17; № 2, стр. 96—103. Совместно с В. Нарбутом.
Оперативное вмешательство при хронической хорее, осложняющей падучую. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900—1901 гг.. СПБ, 1901, стр. 67—69.
Природа гипноза. Вестник знания, 1926, № 1, стр. 35—40.
Самовнушение и куеизм, как исцеляющий фактор. Вестник знания, 1925, № 17—18, стр. 1121 — 1129.
Светолечение в нервных болезнях. Обозрение психиатрии, неврологии и экспериментальной психологии, 1916, № 1—3, стр.67—89.
Случай навязчивых идей, излеченный самовнушениями в начальных периодах гипноза. Вестник клинической и судебной психиатрии и невропатологии, 1890, в. 2, стр. 153—166.
Случай операции при эпилепсии. Врач, 1895, № 17, стр. 488.
Хирургия при душевных болезнях. Обозрение психиатрии, неврологии и экспериментальной психологии, 1908, № 2, стр. 79— 94; № 4, стр. 208—227. Совместно с Л. М. Пуссепом.
Выступления в прениях по поводу сообщений
Агаджаияиц К- С., О заграничной командировке по озиаком. лению с препаратом Эрли-ха «606». Протоколы заседаний Общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр. 68— 69 и 76—77.
Аствацатуров М. И., О применении Vier-Zellen-Bad в терапии нервных болезней. Отчеты научных собраний врачей
С.-Петербургской клиники душевных и нервных болезней за 1905—1906 и 1906—1907 гг., СПБ, 1908, стр. 34. Боришпольский Е. С, Лечение седалищной боли шотландскими душами. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 и 1896—1897 гг., СПБ, 1898, стр. 45.
Боришпол ь с к и й, Е. С, О влиянии дрожания на возбудимость коры и мозговое кровообращение. Отчеты научных собраний вра-. чей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 и 1896—1897 гг., СПБ, 1898, стр. 95.
ВалицкаяМ. К., Демонстрация аппарата для парализованных. Протоколы заседаний Общества психиатров за 1899 г., СПБ, 1901, стр. 21.
Горелов А. А., Предметное и словесное внушение в бодрствен-иом состоянии у здоровых и у лиц, страдающих неврастенией и истерией. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—1909 и 1909— 1910 гг., СПБ, 1910, стр. 125.
Грибоедов А. С, Лечение невралгий электрическим светом. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898—1899 гг., СПБ, стр. 62,
Жуков Н. А., Продолжительная электризация мышц постоянным током и ее влияние на мышечную силу. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 и 1896— 1897 гг., СПБ, 1898, стр. И.
Израэльсон Ж. И., К вопросу о механизме возникновения навязчивых состояний и к технике психоанализа. Протоколы заседаний Общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр. 95— 96.
Карпинский А. И., Лечение бессонницы. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных
511
болезней за 1899—1900 гг., СПБ, 1900, стр. 70—71.
Н и к и т и н М. П., Материалы к вопросу о лучах Blondlot и Char-pentier. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1904—1905 гг., СПБ, 1905, стр. 25.
Ольдерогге В. В., Клинические наблюдения над действием ско-поламина при эпилепсии. Протоколы заседаний общества психиатров за 1899 г., СПБ, 1901, стр. 18.
Ольдероге В.В. и Юрман Н Л.О действии скополамина на душевнобольных. Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 95—96.
Оморокова Л. И., Влияние интеркуриругощих инфекционных заболеваний на течение и исход психозов. Протоколы заседаний Общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр. 13—14.
Осипов В. П., О влиянии опо-церебрина на возбудимость мозговой коры. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902—1903 гг., СПБ, 1903, стр. 12—13.
Певиицкий А. А., Лечение алкоголиков гипнозом в амбу-лансе клиники академика В. М. Бехтерева. Отчеты научных собраний врачей С.-Петербургской клиники,, душевных и нервных болезней за 1902— 1903 гг., СПБ, 114—115.
Пуссеп Л. М., Влияние препарата «бОб1» на сифилитические и парасифилитические заболевания центральной нервной системы по собственным наблюдениям. Протоколы заседаний Общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр. 84—85.
Пуссеп Л. М., Демонстрация больного с иссечением гассерова узла. Протоколы заседания Общества психиатров за 1909 и 1910 гг., СПБ, 1911, стр. 78—79.
Пуссеп Л. М., Светолечение в Париже, Гамбурге и Берлине. Отчеты научных собраний вра-
чей С.-Петербургской клиники душевных и нервных болезней за 1900—1901 гг., СПБ, 1901, стр. 16.
Пуссеп Л. М. Случай поражений кожи после освещения радием. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1903—1904 гг., СПБ, 1904, стр. 11.
Пуссеп Л. М., Экспериментальные данные о влиянии церебри-на на мозговое кровообращение. Протоколы заседаний Общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 26.
Рыбаков Ф. Е. и Розен-т а л ь А. Г., Лечение алкоголиков. Труды II съезда Отечественных психиатров в 1905 г., Киев, 1907, стр. 341.
С о л у х И. П., Случай ticconvulsif, излеченный светом вольтовой дуги. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899—1900 гг., СПБ, стр. 65.
Суханов С. А., Современное учение о психотерапии. Вестник психологии, криминальной антропологии и гипнотизма, 1910, № 2, стр. 199.
ТекутьевФ. С, К лечению эпилепсии Adonis vernalis. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 гг. и 1896—1897 гг., СПБ, 1898, стр. 89—91.
ТекутьевФ. С,О лечении больного эпилепсией. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897— 1898 гг., СПБ, 1899, стр. 26—27.
Т у в и м Р. И., Клинические наблюдения над лечением белой горячки. Протоколы заседаний Общества психиатров за 1898 г., СПБ, 1899, стр. 18.
Шайкевич М., О лечении хореи большими дозами мышьяка. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902—1903 гг., СПБ, 1903, стр. 118—120.
512
IV. ПСИХОЛОГИЯ И ПЕДАГОГИКА
Общие вопросы
Книги
Бессмертие человеческой личности как научная проблема. Пг., 1918, 24 стр.
Внушение и его роль в общественной жизни, СПБ, 1903, 144 стр. То же, СПБ, 1908, 175 стр.
Психика и жизнь, СПБ, 1902, 138 стр. То же, СПБ, 1904, 206 стр.
Психология, рефлексология и марксизм, Л., 1925, 80 стр. Часть тиража вышла с заглавием «Психология или рефлексология».
Роль внушения в общественной жизни. Обозрение психиатрии, неврологии и экспериментальной психологии, 1898, № 1, стр. 1 — 19; № 2, стр. 81—99; № 3, стр. 161—177. То же, отд. изд., СПБ, 1898, 55 стр.
Статьи
Биологическое значение психики. Вестник психологии, криминальной антропологии и гипнотизма, 1904, в. 1, стр. 1 —11; в. 2, стр. 83—89 в. 3, стр. 114— 123.
Внушение как фактор преступлений. Обозрение психиатрии, неврологии и экспериментальной психологии, 1899, № 9, стр. 748— 749.
Личность и труд. В кн.: М ю н с-терберг, Бехтерев и М еде, Сборник статей по прикладной психологии, Гос-техиздат, 1922, стр. 62—90.
Мысленное внушение или фокус? Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 8, стр. 584— 585.
О зоорефлексологии как научной дисциплине и о разговоре попугаев с точки зрения объективного, resp. рефлексологического, исследования. В кн.: Вопросы изучения и воспитания личности, 1921, в. 3, стр. 453—474; в. 4—5, стр. 747—804.
Отличном и общем сознании. Вестник психологии, криминальной антропологии и гипнотизма,
1904, в. 8, стр. 541—552; в. 9, стр. 625—641.
Об опытах над «мысленным* воздействием на поведение животных. Вопросы изучения и воспитания личности, 1920 в. 2, стр. 230—265.
Об основных законах мира при объективном рассмотрении соотносительной деятельности человека и его социальной жизни с точки зрения рефлексологии. В кн.: Вопросы изучения и воспитания личности, 1920, в. 2, стр. 171 — 204; 1921, в. 3, стр. 353-396; 1922, в. 4—5, стр. 501—530.
Об отношении рефлексологии и социологии. Обозрение психиатрии, неврологии и рефлексологии, 1926, № 3, стр. 213—216.
Общие итоги деятельности II съезда но педагогической психологии. Заключительное слово ■ при закрытии съезда. Вестник психологии, Криминальной антропологии и гипнотизма, 1910, № 4, стр. 225—230.
Одаренность, гениальность и мозг. Вестник знания, 1927, № 14, стр. 834—886.
Предмет и задачи общественной психологии как объективной науки. Вестник знания, 1911, № 1, стр. 17—24. То же, отд. отт., СПБ, 1911, 19 стр.
Природа сновидений. Вестник знания, 1926, № 22, стр. 1425— 1432.
Роль внушения в общественной жизни. Обозрение психиатрии, неврологии и рефлексологии, 1928, № 1, стр. 1 — 19; Н« 2, стр. 81—99; № 3, стр. 161 — 177.
Сознание и его границы. В кн.: Годичный акт в Казанском университете 5 ноября 1888 г., Казань, 1888, стр. 1—32. То же, отд. отт., Казань, 1888, 82 стр. Реф. (П. И. Ковалевский). Архив психиатрии, неврологии и судебной психопатологии, 1889, № 2, стр. 106.
Что такое внушение? Вестник психологии, криминальной антропологии и гипнотизма, 1904, в. 1, стр. 35—49.
33 В. М. Беетерег,
Г) 13
Экспериментальные исследования над выражением душевных движений у животных. Врач, 1884, № 1, стр. 1—4.
Выступления в прениях но поводу сообщений
Л а иду Л. А., Экспериментальное исследование о памяти мышечного чувства. Протоколы заседаний Общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 102.
Ф а л ь к М. Л., Опыты над памятью активных движений. Протоколы заседаний Общества психиатров за 1897 г., СПБ, 1899, стр. 59.
Ш и л о в Н. Н., Психология женщины и мужчины. Протоколы заседаний Общества психиатров за 1899 г., СПБ, 1901, стр.12—13.
Шумков Г. Е., О воспроизведении активно-двигательных раздражений. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—1909 и 1909 1910 гг., СПБ, 1910, стр. 32, 42—43.
Щеглов Ю. Л., Экспериментально-психологическое исследование но вопросу о внушаемости малолетних преступников. Протоколы заседаний Общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 108.
Объективная психология и рефлексология
Книги
Биологическое развитие мимики с объективно - психологической точки зрения, СПБ, 1910, 38 стр.
Задачи и метод объективной психологии, СПБ, 1909, 12 стр.
Значение органов равновесия в образовании представлений о пространстве, СПБ, 1896, 52 стр. То же, отд. изд., СПБ, 1898, 52 стр.
Коллективная рефлексология, Пг., 1921, 432 стр.
Общие основания рефлексологии, Пг., 1918, 163 стр.
Общие основы рефлексологии человека. М.—Пг., 1923, 408 стр.
То же, Л., 1926, 423 стр. (библиогра-
фия стр. 411 —418). То же, М.—Л., 1928, 544 стр. (библиография стр. 529—537).
Объективная психология (специальная часть). Вестник психологии, криминальной антропологии и гипнотизма, 1910, в. 2, стр. 157— 186; в. 3, стр. 205—244; в. 4, стр. 309—351; в. 5, стр. 467— 502.
Объективная психология, СПБ, 1907—1910, вв. 1—3, 660 стр.
Объективное изучение личности, Пг.г 1923, в. 1, 63 стр.
Объективно-психологический метод в применении к изучению преступности, СПБ, 1912, 61 стр. То же в кн.: Д. А. Д р и л ь, Учение о преступности и мерах борьбы с нею, СПБ, 1912, стр. 507— 568.
Работа головного мозга (с рефлексологической точки зрения), Л., 1926, 93 стр.
Рефлексология труда. М.—Л., 1926, 188 стр. с ил л. Совместно с Л. Л. Васильевым и А. Ф. Вербовым.
Теория образования наших представлений о пространстве. Вестник клинической и судебной психиатрии и невропатологии, 1884, в. 1, стр. 206—252. То же, отд. изд., СПБ, 1884, 52 стр.
Статьи
Данные эксперимента в области коллективной рефлексологии. В сб.: Новое в рефлексологии и физиологии нервной системы, 1925, ' в. 1, стр. 306—337.
Демонстрация новых таблиц для объективного исследования некоторых нервно-психических функций. Обозрение психиатрии, неврологии и экспериментальной психологии, 1910, № 7, стр. 439—441. Содокладчик С. С. Владычснко.
Значение исследования двигательной сферы для объективного изучения нервно-психической деятельности человека. Русский врач, 1909, № 33, стр. 1105— 1109—11 № 35, стр. 1171—1176; № 36, стр. 1199—1203. Реф. (С. Доброгаева): Обозрение психиатрии, неврологии и экспериментальной психологии, 1910, № 3, стр. 171 — 173.
Г: 1 -1
Значение эксперимента в коллективной рефлексологии. В кн.: Научные известия, ст. 1, М., 1922, стр. 34—50.
Личность художника в рефлексологическом изучении. «Арена», «Театральный альманах», Пг., изд. «Время», 1924, стр. 25—44.
Личные рефлексы. Вестник психологии, криминальной антропологии и гипнотизма, 1912, т. 9, в. 1, стр. 1—49.
Мимико-соматический рефлекс на-стораживания. В кн.: Новое в рефлексологии и физиологии нервной системы, в. 1, 1925, стр. 249—253. Автореферат. Совместно с Г. Е. Шумковым.
Мимические рефлексы. Вестник психологии, криминальной антропологии и гипнотизма, 1911, в. 1, стр. 1—48.
Новый аппарат для исследования слухового восприятия. Вестник психологии, криминальной антропологии и гипнотизма, 1904, в. 7, стр. 500—504.
О биологическом развитии человеческой речи. Вестник психологии, криминальной антропологии и гипнотизма, 1911, в. 2, стр. 73—86.
О значении органов равновесия в отношении развития наших представлений о пространстве. Неврологический вестник, 1895, т. 3 в. 4, стр. 107—154.
О взаимоотношении различных двигательных сочетательных рефлексов. Обозрение психиатрии, неврологии и экспериментальной психологии, 1914'—1915, № 10, 11, 12, стр. 543—547.
О влиянии гипноза и внушения на сочетательные рефлексы. В сб.: Новое в рефлексологии и физиологии нервной системы, в. 1, 1925, стр. 179—204. Совместно с Н. М. Щеловановым. .О методах изучения репродуктивной сочетательной реакции и сфере движения у человека. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907—1909 гг., СБП, 1909, стр. 69—70.
О методах объективной психологии. Обозрение психиатрии, невро-
логии и экспериментальной психологии, 1908, № 1, стр. 50—51.
О причинах обмолвок речи. Голос п речь, 1913, № 9, стр. 3—6.
О репродуктивной сочетательной реакции в движениях. Обозрение психиатрии, неврологии и экспериментальной психологин, 1908, № 7, стр. 385—389.
О рефлексах сосредоточения. Вестник психологии, криминальной
антропологии и гипнотизма, 1911, в. 3, стр. 1—44.
О соотношении объективных проявлений сочетательных рефлексов с субъективным показанием исследуемых лиц, Обозрение психиатрии, неврологии и экспериментальной психологии, 1914— 1915, № 10, 11, 12, стр. 558— 562.
О творчестве с рефлексологической точки зрения. В кн.: Грузенберг С. О. Гений и творчество, Л., 1924, стр. 228—233.
Об общих основах рефлексологии как научной дисциплины. Природа, 1917, № 11, 12, октябрь-ноябрь, стр. 1101 — 1128.
Об основных проявлениях нервно-психической деятельности в объективном их изучении. Русский врач, 1911, № 12, стр. 457—463.
Об основах функциональной деятельности мозговой коры но данным нсихорефлексологии. Русский врач, 1913, № 33, стр. 1149—1155.
Об экспериментальном психологическом исследовании преступников. Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 8, стр. 568—570.
Об эмоциях как мимико-соматиче-ских рефлексах. В кн.: Новое в рефлексологии и физиологии нервной системы, Сборник 3, посвященный памяти В. М. Бехтерева, М.—Л., 1929, стр. 17—30.
Обоснование объективной психологии. Вестник психологии, криминальной антропологии и гипнотизма, 1907, в. 1, стр. 1 —17, в. Ill, стр. 89—127.
Образование наших представлений о пространстве. Протоколы заседаний общества психиатров
33* 515
в. С.-Петербурге в 1884 г., СПБ, 1885, стр. 29—34.
Объективное исследование нервно-психической деятельности. Обозрение психиатрии, неврологии и экспериментальной психологии, 1907, № 9, стр. 513—536.
Объективная психология. Новое слово, 1909, № 6, стр. 28—34.
Объективная психология и ее предмет. Вестник психологии, криминальной антропологии и гипнотизма, 1904, № 9, стр. 650— 661; № 10, стр. 721—736.
'Основные задачи рефлексологии физического труда. В кн: Вопросы изучения и воспитания личности. Пг, 1920, в. 1, стр. 3—51.
Основные принципы так называемой объективной психологии или психорефлексологии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1910, № 10—11, стр. 577—595.
Основные принципы работы мозговой коры. Сборник в честь пятидесятилетия врачебно-об-щественной и научио-педагоги-ческой деятельности проф. В. Я- Данилевского, 1925, стр. 1 — 11.
Перспективы рефлексологии человека. Вестник знания, 1927, № 6, стр. 320—326.
Половая деятельность с точки зрения рефлексологии. В сб.: Половой вопрос в свете научного знания, М.—Л., 1926, стр. 142— 182.
Принцип скачкообразного развития сочетательно-рефлекторной деятельности. В кн.: Вопросы изучения и воспитания личности, 1920, № 1, стр. 69—87; № 2—3, стр. 115—158. Совместно с Я- Г. Полонским.
Рефлекс настораживания или подготовки. Современная психоневрология, 1926, № I—2, стр. 7—12; № 3, стр. 115; № 4, стр. 229—238. Совместно с Г. Е. Шумковым.
Рефлексы сосредоточения. Вестник психологии, криминальной антропологии и гипнотизма, 1911, в. 3, стр. 1—44.
Роль сосредоточения как доминанты в процессах сочетательно-рефлекторной деятельности. Вопросы изучения и воспитания
личности, 1927, № 1—2, стр. 3—18.
Символические рефлексы. Вестник психологии, криминальной антропологии и гипнотизма, 1911, в. 4, стр. 1—62; в. 5, стр. 1—34.
Строго-объективный метод в изучении нервно-психической, resp. соотносительной, деятельности и его роли в обосновании рефлексологии человека. Вестиик знания, 1917, № 4—5, стр. 228— 245.
Что такое объективная психология. Вопросы философии и психологии, 1909, кн. IV, стр. 696— 703.
Выступления
в прениях по поводу
сообщений
Анфимов В. Я., Демонстрация нового типа таблиц букв для исследования внимания. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1908—1909 и 1909—1910 гг., СПБ, 1910, стр. 36.
Владимирский А. В., О скорости умственных процессов у глухонемых. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907—-1908 гг., СПБ, 1909, стр. 47.
Грибоедов А. С, Влияние цветного освещения на память слов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897—1898 гг., СПБ, 1899, стр. 67.
Д о б р о т в о р с к и й Н. М., О сочетательной реакции в двигательной сфере. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907— 1908 гг., СПБ, 1909, стр. 71—72.
Лазурский А. Ф-, Влияние представления иа ход ассоциаций. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898—1899 гг., СПБ, 1899, стр. 41.
Лазурский А. Ф., К вопросу об экспериментальном анализе
516
апперцепции. Вестник психологии, криминальной антропологии и гипнотизма, 1910, Л1» 4, стр. 429.
Лазурский А. Ф., О влиянии мышечных движений (ходьбы) на скорость умственных процессов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 и 1896— 1897 гг., СПБ, 1898, стр. 68.
Лазурский А. Ф., О влиянии различного чтения на ход ассоциаций. Протоколы заседаний Общества психиатров за 1899 г., СПБ, 1901, стр. 45.
Лазурский А. Ф. иШико-в а Н. Н., Память последовательных зрительных впечатле-: ний. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1898—1899 гг., СПБ, 1899, стр. 76—77.
Л и щ и н а Н. Г., Исследование колебаний внимания при восприятии однородных зрительных впечатлений. Вестник психологии, криминальной антропологии и гипнотизма, 1904, № 2, стр. 87.
Никитин М. П., К вопросу об образовании зрительных восприятий (экспериментальные исследования). Вестник психологии, криминальной антропологии и гипнотизма, 1904, № 3, стр. 183.
Павловская Л. С. и Еланская В . Н., Экспериментальное исследование способности суждения. Вестник психологии, криминальной антропологии и гипнотизма, 1904, № 1, стр. 82.
Поварнин К. И., Об условиях привлечения и сосредоточения внимания. Вестник психологии, криминальной антропологии и гипнотизма, 1904, № 8, стр. 619.
Телятник Ф. К., О зритель-нон иллюзии Muller-Lyer'a. От четы научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 и 1896—1897 гг., ' СПБ, 1898, стр. 26—27.
Щеглов А. Л., Экспериментально-психологические исследова-
ния по вопросу о работоспособности малолетних преступников. Вестник психологии, криминальной антропологии и гипнотизма, 1904, № 1, стр. 82—83.
Психология развития и проблемы воспитания
Книги
Внушение и воспитание, СПБ, 1912, 20 стр. То же, Пг., 1928, стр. 40.
Вопросы воспитания в возрасте первого детства. Образование, 1909, № 2, стр. 24—69. То же, отд. отт., СПБ, 1909, стр. 39.
Вопросы общественного воспитания. Вестник воспитания, 1909, № 9, стр. 103—141. То же, отд. изд., М., 1910, 41 стр.
Вопросы эволюции нервно-психической деятельности и отношение к педагогике. Вестник психологии, криминальной антропологии и гипнотизма, 1916, в. 1, стр. 1 — 16. То же, отд. изд., Пг., 1916, 16. стр.
Значение музыки в эстетической воспитании ребенка с первых дней его детства. Вестник воспитания, 1915, № 6, стр. 47—57. То же, отд. изд., М., 1916, 13.стр.
О развитии нервно-психической деятельности в течение первого полугодия жизни ребенка. Вестник психологии, криминальной антропологии и гипнотизма, 1912, в. 2, стр. 1—48. То же, отд. изд., СПБ, 1912, 48. стр.
О социальнотрудовом воспитании, Пг., 1917, 16 стр.
Об индивидуальном развитии нервно-психической сферы по данным объективной психологии. Вестник психологии, криминальной антропологии и гипнотизма, 1910, № 11, стр. 105—121. То же, отд. изд., СПБ, 1910, 16 стр.
Объективное исследование нервно-психической сферы в младенческом возрасте, СПБ, 1909, 18 стр.
Охрана детского здоровья. К вопросу о воспитании детей дошкольного возраста, СПБ, 1911, 24 стр.
Первоначальная эволюция детского рисунка в объективном изучении. Вестник психологии, кри-
517
минальной антропологии и гипнотизма, 1910, в, 1., стр. 1—50. То, же, отд. изд., СПБ, 1910, 50 стр.
Статьи
К обоснованию генетической рефлексологии. В сб.: Новое в рефлексологии и физиологии нервной системы, в. 1, 1925, стр. 116—132.
Культура социального героизма в воспитании детей. Вестник знания, 1926, № 24, стр. 1571 — 1574.
Общие вопросы
Книги
Личность и условия ее развития И здоровья. Речь, произнесенная 4' сентября 1905 г. на II съезде отечественных психиатров, СПБ, 1905, 43 стр.
Статьи
Борьба за науку. Вестник знания, 1925, № 14, стр. 929—932.
Вопросы нервно-психического здоровья в русском населении. Обозрение психиатрии, неврологии и экспериментальной пснхо-. логии, 1910, № 6, стр. 321—343.
Значение Ломброзо в вопросе о , преступности. Запросы жизии, 1909,' № 9, стр. 8—11.
Индивидуальные и социальные факторы развития организмов и социальность как условие прогресса. Вестник психологии, криминальной антропологии и гипнотизма, 1914, в. 1, стр. 1 — 18. Личность, условия ее умственного и нравственного развития. Обозрение психиатрии, неврологии и экспериментальной психологии, 1905, № 8—9, стр. 630—645.
Недооценка социальной роли женщины. Вестник знания, 1925, № 11, стр. 771—774.
О причинах самоубийства и о возможной борьбе с ннм. Вестник знания, 1912, № 2, стр. 131 — 141; № 3, стр. 253—264.
Письмо по случаю 30-летия клиники
О воспитании в младенческом возрасте. Вестник психологии, криминальной антропологии и гипнотизма, 1913, в. 3, стр. 1;—28. То же, отд. изд., СПБ, 1913, 20 стр.
Об объективном исследовании детской психики. Вестник психологии, криминальной антропологии и гипнотизма, 1908, в. 1, стр. 4—17.
Об эволюции нервно-психической деятельности. Русский врач, 1913, № 14, стр. 457—462; № 15, стр. 497—500
душевных и нервных болезней. Военно-медицинская академия. Неврологический вестник, 1897, т. 5, № 4, стр. 217—219.
Приветственное слово, сказанное при открытии первого съезда по борьбе с. пьянством. Обозрение психиатрии, неврологии и экспериментальной психологин, 1910, № 5, стр. 257—258.
Первые шаги культурного и научного сближения славян. Вестник знания, 1909, №2, стр. 173— 185; № 3, 336 стр.
Проблески будущего (по поводу субботников . Железнодорожная техника и Зкономика, 1919, № 3, 202 стр.
Протест по поводу продовольственной блокады. Петроградская правда, 1920 от 1 января.
Социальное бессмертие человеческой личности. Вестник знания, 1928, № 1, стр. 4—П.
Человек железной воли. Рассказ об участии в лечении В. И. Ленина в Горках, Петроградская правда, 1924 от 26 января, № 21.
Что дала инициативная конференция по научной организации труда. В сб.: Вопросы изучения труда. Пг., 1922, стр. 5—26
Выступления в прениях по поводу сообщений
Б у р и н с к и й Е. Ф., Уранизм как социальное явление. Отчеты научных собраний врачей
V ОБЩЕСТВЕННЫЕ ВЫСТУПЛЕНИЯ И ОРГАНИЗАЦИОННАЯ ПСИХОНЕВРОЛОГИЯ
Г> 18
С-ПетерОургской клиники душевных и нервных болезней за 1902—1903 гг., СПБ, 1903, стр. 81—84.
В я ш к о в В. М., К статистике душевных заболеваний в крестьянском населении С.-Петербургской губернии. Протоколы заседаний общества психиатров за 1895—1896 гг., СПБ, 1897, стр. 92—93.
И г и а т ь е в М. В., Данные статистики душевнобольных в России. Протоколы заседаний общества психиатров за 1894 г., СПБ, 1896, стр. 20.
И I' на т ь е в М. В., О причинах душевных болезней по данным отчетов русских больниц. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1900—1901 гг., СПБ, 1901, стр. 53—54.
Лебедев И. П., О покушении на самоубийство с медицинской точки зрения. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 и 1896—1897 гг., СПБ, 1898, стр 15—16.
Островских, Нервные и душевные заболевания в Персии. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1897—1898 гг., СПБ, 1899, стр. 39—40.
Психогигиена и психопрофилактика
Книги
Алкоголизм и борьба с ним, Л.,
1927, 62 стр.
Значение полового влечения в жизнедеятельности организма, М.,
1928, 32 стр.
Об алкогольном оздоровлении. В кн.: Вопросы алкоголизма, СПБ, 1913, стр. 81 — 100. То же, отд. отт., СПБ, 1912, 4 стр.
Существенное в вопросе о борьбе
с алкоголизмом. Вестник знания, 1912, № 10, стр. 787—791. То же, отд. отт., СПБ, 1912, 4 стр.
Статьи
Алкоголь и эпилепсия. В кн.: Вопросы алкоголизма, СПБ, 1913,
стр. 101 —120. Совместно "с В. Я. Анфимовым.
Алкогольная политика и алкогольное оздоровление. Вестник Европы, 1912, № Ю, стр. 290— 298. То же, отд. изд., СПБ, 1912, 11 стр.
Вопросы'алкоголизма н меры борьбы с его развитием. Вопросы алкоголизма, СПБ, 1913, стр. 13—33. То же, Русский врач, 1912, № 22, стр. 951—957.
Воспитание умственной активности. Вестник знания, 1928, № 23— 24, стр. 1090—1091.
К алкогольному вопросу. Врачебная газета, 1912, № 43, стр. 1540—1541.
О половом оздоровлении. Вестник знания, 1910, № 9, стр. 924— 935; № 10, стр. 1043—1050.
Об алкогольном оздоровлении. Русский врач, 1912, № 36, стр. 1449—1456.
Отрезвление народа и физическое и нравственное его оздоровление. Русский врач, 1915, № 15, стр-. 342—345.
Рациональное использование человеческой энергии в труде. Труды 1-й Всероссийской инициативной конференции по научной организации труда и производства, в. 1, М., 1921, стр. 23— 34.
Социальное наследие старого быта. Вестник знания, 1927, № 9, стр. 531—538.
В ытс туплен ия в прениях п_о поводу сообщений
Р е и т ц Г. В., О влиянии хронического алкоголизма иа развитие организма. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1899— 1900'гг., СПБ, 1900, стр. 29.
Организация научных обществ, учебных заведений, исследовательских учреждений
Статьи
Задачи психо-неврологического института. Речь,произнесенная при открытии курсов института, февраль 1908. Вестник знания, 1908, № 3, стр. 3—19.
519
Г.ще несколько слов по поводу проектируемого русского союза психиатров и невропатологов. Обозрение психиатрии неврологии и экспериментальной психологии, 1902, № 6, стр.471 — 478.
К вопросу о первоначальном возникновении Союза психиатров и невропатологов. Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 3, стр. 233.
К предварительному проекту устава русского союза психиатров и невропатологов. Обозрение психиатрии, 1896, № 10, стр. 765— 775.
О деятельности Психоневрологического института как высшего педагогического учреждения и его роли в педагогическом строительстве. Вопросы изучения и воспитания личности, в. 3, Пг., 1921, стр. 435—452.
О славянском съезде врачей в г. Софии. Русский врач, 1910, № 33, стр. 1175—1177.
О современном развитии неврологических знаний и значении в этом развитии научных обществ. Неврологический вестник, 1893, т. 1, в. 1, стр. 1 —14. То же, отт., Казань, 1893, 16 стр.
Об организации славянского научного комитета при Психоневрологическом институте. Вестник знания, 1909, № 5, стр. 614—616.
Об устройстве Психоневрологического института в С.-Петербурге. Вестник психологии, криминальной антропологии и гипнотизма, 1904, в. 5, стр. 273—276.
Об учреждении Русского союза психиатров и невропатологов. Обозрение психиатрии,неврологии и экспериментальной психологии, 1897, № 3, стр. 177—179.
От редакции. Обозрение психиатрии, неврологии и рефлексологии, 1926, в. 1, стр. 1—2. За подписью В. М. Бехтерева сообщается о перемене в заголовке журнала при его возобновлении в 1926 г.
Пантеон мозга. Вестник знания, 1927, № 21, стр. 1315—1318.
По поводу открытия Союза отечественных психиатров. Обозрение психиатрии, неврологии и экс-
периментальной психологии
1910, № 5, стр. 279—281.
По поводу учреждения Русского •союза психиатров и невропатологов. Обозрение психиатрии, неврологии и экспериментальной психологии, 1902, № 7 стр. 544—553.
Предположение об учреждении Психоневрологического института в С.-Петербурге. Вестник психологии, криминальной антропологии и гипнотизма, 1904, в. 5, стр. 357.
Речь при открытии научных собраний врачей при клинике нервных и душевных болезней. Врач, 1895, № 42, стр. 1189—1190.
Выступления
Высказывание по поводу истории возникновения Союза психиатров и невропатологов. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 66— 67, 70.
Выступление по поводу IX Пироговского съезда. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 —1902 гг., СПБ, 1902, стр. 70—71.
Выступление по поводу событий в больнице Николая Чудотворца. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1905—1906 и 1906—1907 гг., СПБ, 1908, стр. 9, 11.
Выступление по поводу устава Общества психиатров. Протоколы заседаний Общества психиатров за 1897 г., СПБ, 1899, стр. 23—24
Организация психиатрической
и невропатологической
помощи
Статьи
Вступительная речь на съезде Общества русских врачей в память Н. И. Пирогова в СПБ с 4—11 января 1904 г. (Секция душевных и нервных болезней 5 января). Обозрение психиатрии, неврологии и экспериментальной психологии, 1904, № 1, стр. 70—74.
520
О кандалах у испытуемых душевнобольных арестантов. Обозрение психиатрии, неврологии и экспериментальной психологии, ■ 1910, № 4, стр. 202—211. Совместно с П. А. Останковым.
О постельном содержании душевнобольных. Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 1, стр. 1—6. Реф.: Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 и 1896—1897 гг., СПБ, 1898, стр. 62—64.
О привлечении частной благотворительной помощи к вопросу о призрении душевно- и нервнобольных и об организации и задачах попечительств о душевио-и нервнобольных, СПБ, без обозначения года, без титульного листа и без обложки, 12 стр.
Об устройстве клиники для больных страдающих последствиями алкоголизма. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1910—1911 гг., СПБ, 1911, стр. 16—18.
Об устройстве соединенных открытых и закрытых заведений для душевно- и нервнобольных. Обозрение психиатрии, неврологии и экспериментальной • психологии, 1901, № 1, стр. 8—9.
Открытие новой клиники нервных болезней. Речь, произнесенная при открытии клиники нервных болезней 19 ноября 1897 г., СПБ, 1897, 10 стр. То же. Обозрение психиатрии, неврологии и экспериментальной психологии, 1897, № 11, стр. 801—818.
Отчет о деятельности института для нервных детей имени акад. В. М. Бехтерева Психоневрологической академии. В кн.: Вопросы из\чения и воспитания личности, 1926, № 2—3, стр. 231—234.
Предположение относительно регистрации состояния больных. От-
четы научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1907—1908 гг., СПБ, 1909, стр. 8—9.
Выступления в прениях по поводу сообщений
Левчаткин В. И., О постельном содержании больных как терапевтическом методе. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895— 1896 и 1896—1897 гг., СПБ,
1898, стр. 24—25. Малеев В. П., О состоянии больницы св. Николая Чудотворца за 1901 г. Протоколы заседаний общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 100, 101, 102, 104.
Осипов В. П., Постельное содержание в женском отделении клиники душевных болезней. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1895—1896 н 1896—1897 гг., СПБ, 1898, стр. 101—102.
Останков П. А., О возникновении системы no-restraint в России. Протоколы заседаний общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 45, 46.
Топорков Н. Н., О необходимости борьбы с туберкулезом в лечебницах душевнобольных. Протоколы заседаний Общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 83.
Шайкевич М. О., Настоящее и желательное будущее отделений для нервно-душевнобольных при военных госпиталях. Труды 11 съезда отечественных психиатров в 1905 г., 1907, стр. 247.
Ю р м а н Н. А., Женский уход за* душевнобольными мужчинами. Протоколы заседаний общества психиатров за 1898 г., СПБ,
1899, стр. 27—28, 29.
VI. ВОСПОМИНАНИЯ, КОРРЕСПОНДЕНЦИИ, ЗАМЕТКИ И СТАТЬИ ПО РАЗЛИЧНЫМ ВОПРОСАМ
Автобиография (посмертная), М.,
1928, 51 стр. Автобиография. Владимир Михлй.ю
вич Бехтерев. Огонек, 1927' № 35. Братство народов и Лев Толстой.
521
Вестник знания, 1925, № 1, стр. 6—10.
Вотяки, их история и современное состояние. Вестник Европы, 1880, т. 4, стр. 621—654; т. 5, стр. 141 — 172.
Выступление В. М. Бехтерева об увековечении памяти И. М. Ба-лииского. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1901 — 1902 гг., СПБ, 1902, стр. 61—63.
Выступление по поводу решения Общества психиатров в С.-Петербурге об увековечивании памяти И. М. Балииского. Протоколы заседаний общества психиатров за 1902 и 1903 гг., СПБ, 1904, стр. 28—31.
Выступление в честь И. И. Мечникова на соединенном заседании С.-Петербургских медицинских обществ 14 мая 1909 г. Обозрение психиатрии, неврологии и экспериментальной психологии, 1909, № 5, стр. 319.
Из воспоминаний о Шарко. Вестник знания, 1926, № 12, стр. 829— 830.
К воспоминаниям о Шарко. Современная психоневрология,. 1925, № 8, стр. 14—17.
Корреспонденции с театра военных действий во время русско-турецкой войны. Северный вестник (газета), 1877, № 55, 24/VI, стр. 3; 1/VII, № 62, стр. 3; 4/VII, № 65, стр. 2; 6/VII, № 67, стр. 2—3; 9/VII, № 70, стр. 2—3; 20/V1I, № 81, стр. 2; 24/VII, № 85, стр. 4; 25/VII, № 86, стр. 2; 27/VII, № 88, стр. 2; 9/VIII, № 101, стр. 3; 10/VIII, № 102, стр. 2—3; 14/VIII, № 106, стр. 3; 24/VIII, № 116, стр. 2—3; 25/VIII, № 117, стр. 3; 28/VIII, № 120, стр. 3; 7/1X, № 129, стр. 3; 8/IX, № 130, стр. 2; 19/IX, № 141, стр. 2; 26/IX, № 148, стр. 2; 27/IX, № 149, стр. 2; 28/IX, № 150, стр. 3.
И. П. Мержеевский и его роль в развитии русской психиатрии. Обозрение психиатрии, неврологии и экспериментальной психологии, 1908, № 3, стр. 131—135.
-Лев Толстой и единение народов. Вестник знания, 1915, № 7,
стр. 436—445. То же, отд. изд Пг., 1915, 12 стр.
Мне отмщение и аз воздам. В кн.: ■ Щит. Сборник под ред. Л. Андреева, М. Горького и Ф. Сологуба, М., 1915, стр. 35—38.
Мог бы жить! По поводу смерти Бергонье. Вестник знания, 1925, № 4, стр. 265—267.
Моральные итоги великой мировой войны. Вестник знания, 1915, № 10—11, стр. 655—678. То же, отд. изд., Пг., 1915, 20 стр.
О роли Гоголя в возрождении русского общества. Вестник психологии, криминальной антропологии и гипнотизма, 1909, в. 3, стр. 254—257.
О сближении славянских народов на почве науки. Вестник Европы, 1909 июль, стр. 314—330.
Памяти А. М. Дохмана. Неврологический вестник, 1893, т. 1, в. 2, стр. 1—2.
Памяти Д. А. Дриля. Вестник психологии, криминальной антропологии и гипнотизма, 1910, в. 4, стр. 307.
Памяти Н. О. Ковалевского. Неврологический вестник, 1893, т. 1, в. 1, стр. 3—4.
Памяти акад. И. П. Мержеевского. Неврологический вестник, 1908, т. 15, в. 1, стр. 1—3.
Памяти И. И. Мечникова. Биржевые ведомости (газета), 1916, 6/VII, стр. 2.
Памяти Л. Н. Толстого. Вестник психологии, криминальной антропологии и гипнотизма, 1910, в. IV, стр. 305—306.
Памяти учителей. Обозрение психиатрии, неврологии и экспериментальной психологии, 1910, № 3, стр. 129—135.
Памяти проф. Шарко. Неврологический вестник, 1893, т. 1, в. 3, стр. IV—VI.
Н. И. Пирогов как научный и общественный деятель. Вестник психологии, криминальной антропологии и гипнотизма, 1910, в. V, стр. 520—523.
Речь В. М. Бехтерева иа праздновании его 25-летнего юбилея 2 мая 1903 г. Отчеты научных собраний врачей С.-Петербургской клиники душевных и нервных болезней за 1902—1903 гг., СПБ, 1903, стр. 100—109.
522
Речь, произнесенная на торжественном научном собрании врачей клиники душевных и нервных болезней 2 мая 1903 г., СПБ, 1903, 12 стр.
Слуга и друг больного рассудком (К столетию со дня рождения И. М. Балинского). Обозрение психиатрии, неврологии и рефлексологии, 1927, № 3, стр. 161 — 164.
Смерть раввина. В кн.: Щит. Сборник под ред. Л. Андреева,
М. Горького, Ф. Сологуба, М.
1915, стр. 175. Софийский съезд. Впечатления из
Болгарии. Вестиик знания, 1910,
№ 12, стр. 1223—1236. Убийство Ющинского и психиатро-
психологическая экспертиза.
Врачебная газета, 1913, № 50,
стр. 1819—1843. То же, отд.
изд., СПБ, 1913, 56 стр. Юбилейные дни в Праге, Пг., 1915,
14 стр.
Редактор Л. К- МИХАЙЛОВ Техн. редактор Ю. С. Бельчикоеа Зав. корректорской Л. М. Голицына Переплет художника Л. С. Эрмана
Т08919. Подписано к печати 17/XI 1953г.
МН-55 Ф. бум. 60х921/16=16,5 бум. л.-
33,0 печ. л.+0,25 п. л. (вкл.) 36,5 уч.-изд.л.
44 200 зн. в 1 п. л. Тираж 9000 экз.
Цена 18 р. 50 к. Переплет 2 р.
Зак. 1158
1-я тип. Трансжелдориздата МПС
ОПЕЧАТКИ
|
Страница |
I Строка I Напечатано |
Следует читать |
|
|
5 |
11 сверху |
Е. В. Адамиком |
Е. В. Адамюком |
|
14 |
19 снизу |
т. е. |
как и |
|
18 |
7 сверху |
симуляции, |
симуляции |
|
26 |
16 снизу |
и opticus |
и п. opticus |
|
34 |
8 снизу |
с точностью |
с положительностью |
|
40 |
1 снизу |
на внутренний наружный |
на внутренний и наружный |
|
65 |
8 сверху |
опсывалось |
описывалось |
|
65 |
1 7 сверху |
боковому |
блоковому |
|
77 |
10 снизу |
subiculeum |
subiculum |
|
99 |
6 снизу |
Геле |
Лере |
|
114 |
4 и 5 сверху |
(от 10 до 13 К-А) |
(от 10 до 13 к. р.) |
|
114 |
6 и 7 сверху |
(от 6 до 8 К—А) |
(от 6 до 8 к. р.) |
|
117 |
7 сверху |
от 4 до 5 К—А |
от 4 до 5 к. р. |
|
125 |
Подстрочное примечание, 4 снизу |
1872 |
1882 |
|
126 |
Подстрочное примечание, I снизу |
международная библиотека |
медицинская библиотека |
|
133 |
15 сверху |
видимом |
невидимом |
|
141 |
Подстрочное примечание, 2 снизу |
' in Wirnam |
in Wien |
|
144 |
I снизу |
приводам и мозжечка |
проводами мозжечка |
|
158 |
17 сверху |
вышеуказанные опыты |
вышеуказанные опыты |
|
|
|
делают существование |
снимают всякий raison |
|
|
|
судорожного центра варо- |
d'etre у судорожного |
|
|
|
лиева моста совершенно невероятным |
центра |
|
183 |
13 снизу |
Один |
«Один |
|
198 |
30 сверху |
— временам |
по временам |
|
205 |
1 0 снизу |
как корешки, дающие нача- |
как соединительнотканные |
|
|
|
ло спинным и шейным нер- |
части позвоночника, |
|
|
|
вам, так и соединительно- |
способствующие его движе- |
|
|
|
тканные части позвоночного |
нию, так и корешки. |
|
|
|
канала |
дающие начало спинным и шейным нервам |
|
207 |
Подстрочное примечание, I снизу |
№ 89 |
№ 82 |
|
237 |
30 снизу |
Студеискнм |
Студентским |
|
255 |
16 снизу |
bicipititis |
bicipitis |
|
257 |
Подстрочное примечание. 1 снизу |
Вестник неврологии |
Неврологический вестник. |
|
260 |
9 снизу |
intraspinalis |
infraspinalis |
|
273 |
10 сверху |
Как верный указатель |
здесь был выяснен, как |
|
|
|
органического паралича |
верный указатель орга- |
|
|
|
был выявлен |
нического паралича |
|
292 |
Подстрочное примечание, 1 снизу |
Bd 81 |
Bd 87 |
|
338 |
17 сверху |
обманом |
обманов |
|
382 |
12 сверху |
Ох, |
«Ох, |
|
407 |
27 сверху |
тонические клоническис |
тонические н клоннческие |
|
437 |
4 снизу |
(Дернова, Ярмоленко) |
(Дернова- Ярмоленко) |
Зак. 1158
Внимание! Сайт является помещением библиотеки. Копирование, сохранение (скачать и сохранить) на жестком диске или иной способ сохранения произведений осуществляются пользователями на свой риск. Все книги в электронном варианте, содержащиеся на сайте «Библиотека svitk.ru», принадлежат своим законным владельцам (авторам, переводчикам, издательствам). Все книги и статьи взяты из открытых источников и размещаются здесь только для ознакомительных целей.
Обязательно покупайте бумажные версии книг, этим вы поддерживаете авторов и издательства, тем самым, помогая выходу новых книг.
Публикация данного документа не преследует за собой никакой коммерческой выгоды. Но такие документы способствуют быстрейшему профессиональному и духовному росту читателей и являются рекламой бумажных изданий таких документов.
Все авторские права сохраняются за правообладателем. Если Вы являетесь автором данного документа и хотите дополнить его или изменить, уточнить реквизиты автора, опубликовать другие документы или возможно вы не желаете, чтобы какой-то из ваших материалов находился в библиотеке, пожалуйста, свяжитесь со мной по e-mail: ktivsvitk@yandex.ru